


Ernst-Jan Eggers1,2፣ አቴ ቫን ደር ቡርት።1፣ Sjaak AW ቫን ሄውስደን2, Michiel E. de Vries@ 1, ሪቻርድ ጂኤፍ ቪሴአር ©2, ክርስቲያን ደብሊውቢ ባኬም©2и & Pim Lindhout1
በድንች ውስጥ የሚገኘው የዘረመል ጥቅም በሄትሮዚጎስ ቴትራፕሎይድ ጂኖም በተመረተው ድንች ተስተጓጉሏል። ድንችን ወደ ዳይፕሎይድ ኢንብሬድ-መስመር ላይ የተመሰረተ ኤፍ1-ድብልቅ ሰብል መቀየር የዘረመል ጥቅምን ለመጨመር ተስፋ ሰጭ መንገድ ነው። የ S-locus inhibitor (Sli) ጂን ወደ ዳይፕሎይድ የድንች ጀርምፕላዝም ማስገባቱ በራሱ የዳበረ ዘርን በብቃት ማመንጨት እና በዚህም የድንች መፈልፈያ መስመሮችን መፍጠር ያስችላል። ስለ ስሊ ሎከስ አወቃቀር እና ተግባር ብዙም የሚታወቅ ነገር የለም። እዚህ ላይ የ Sliን ካርታ ወደ 12.6 ኪ.ባ ክፍተት በክሮሞዞም 12 ላይ በእንደገና ስክሪን አቀራረብ እንገልፃለን። በዚህ የጊዜ ክፍተት ውስጥ ከሚገኙት ሁለት እጩ ጂኖች ውስጥ አንዱ በራሱ ተኳሃኝ በሆኑ መስመሮች ውስጥ ብቻ የሚገኝ ልዩ ቅደም ተከተል ያሳያል. ከራስ ጋር የማይስማማውን የሚቀይር አገላለጽ ቬክተር እንገልፃለን። የጂኖታይፕስ ወደ ራስ-ተኳሃኝ እና SC ጂኖታይፕስ ወደ SI የሚቀይር CRISPR-Cas9 ቬክተር። የ Sli ጂን የኤፍ-ቦክስ ፕሮቲንን በኮድ ያስቀምጣል, እሱም በተለይ ከራስ-ተኳሃኝ ተክሎች የአበባ ዱቄት ውስጥ ይገለጻል. የዚያ ጂን አራማጅ ውስጥ 533 ቢፒ ማስገባት ወደ ተግባር ሚውቴሽን ይመራል፣ ይህም የራስ ብናኝ አለመቀበልን ያሸንፋል።
ድንች በዓለም ላይ በጣም አስፈላጊው የእህል ያልሆነ የምግብ ሰብል ነው። ሆኖም እንደ በቆሎ፣ ሩዝ እና ስንዴ ያሉ ሌሎች የምግብ ሰብሎች በዓመት 1 በመቶ የዘረመል ምርት አግኝተዋል።1በድንች ውስጥ ያለው የዘረመል ጥቅም በጣም አናሳ ነው።2. በአሁኑ ጊዜ አብዛኛው ለንግድ የሚለሙ የድንች ዝርያዎች የሚመነጩት በሄትሮዚጎስ አውቶቴትራፕሎይድ ወላጆች መካከል ካሉ መስቀሎች ነው። በዚህ የመራቢያ ሥርዓት ውስጥ በየመራቢያ ትውልዱ በመቶ ሺዎች የሚቆጠሩ ችግኞች ይፈጠራሉ እና ይጣራሉ ይህም እነዚያን ብርቅዬ ግለሰቦች በዘር ውስጥ ለሚለያዩ በርካታ ባህሪያት ተቀባይነት ያላቸው ባህሪያትን ለመለየት ነው. ለንግድ የድንች ዝርያ ዋጋ ጠቃሚ የሆኑ አንዳንድ ሃምሳ ባህሪያት ስላሉ፣ እነዚህን ባህሪያት የሚቆጣጠሩት ምርጥ አሌሎችን በተለመደው የድንች እርባታ በመጠቀም የማጣመር እድሉ እዚህ ግባ የሚባል አይደለም። በተጨማሪም፣ የጄኔቲክ ታማኝነትን በኋለኛ ማቋረጫ ዘዴዎች በመጠበቅ አዳዲስ ባህሪያትን ወደ ምሑር ዝርያዎች ማስተዋወቅ ያለ ግብረ-ሰዶማዊ የወላጅ መስመሮች የማይቻል ነው። እነዚህን ችግሮች ለመቅረፍ በርካታ ቡድኖች የተዳቀለ መስመር ላይ የተመሰረተ ዳይፕሎይድ ድንች የመራቢያ ፕሮግራሞችን ጀምረዋል።2-5. በነዚህ ፕሮግራሞች ውስጥ የዘር ውርስ የተገኘው በዘር ማደግ በሚፈጠር የወላጅ መስመር ማሻሻያ አማካኝነት በዘር ማዳቀል ወቅት ጎጂ የሆኑ አለርጂዎችን ያለማቋረጥ በመምረጥ እና በተፈጠሩት መስመሮች ውስጥ ጠቃሚ የሆኑ አለርጂዎችን በጀርባ ማቋረጫ ዘዴዎች በመደርደር ነው።6. ሄትሮቲክ F1-ድብልቅ ዘሮችን ለማምረት የወላጅ የተዳቀሉ መስመሮች ይሻገራሉ.
በአብዛኛዎቹ የዲፕሎይድ ድንች ጂኖታይፕስ ውስጥ፣ ዘር ማዳቀል በባለብዙ-አሌሊክ ኤስ-ሎከስ ቁጥጥር ስር ባለው ጋሜቶፊቲክ ራስን አለመጣጣም (ጂኤስአይ) ስርዓት በእጅጉ የተገደበ ነው። ይህ ኤስ-ሎከስ የ S-RNases ዘይቤን ይገልፃል ይህም በስልቱ ውስጥ የራስ የአበባ ዱቄት እድገትን የሚገታ፣ ራስን ማዳበሪያን ይከላከላል።7. የአበባ ዱቄት በሚበቅልበት ጊዜ፣ የአበባ ዱቄት-ኤክስፕረስ ኤስ-ሎከስ ኤፍ-ቦክስ ፕሮቲኖች (ኤስኤልኤፍ) ኤስ-አርናሴስን ይገነዘባሉ እና ወደ ፕሮቲአሶማል መበላሸት መንገድ ያነጣጥራሉ ፣ ይህም የአበባ ዱቄት ቱቦ ማዳበሪያ ወደሚገኝበት ኦቫሪ እንዲበቅል ያስችለዋል።8. እያንዳንዱ S-allele S-RNaseን እና በርካታ ኤስኤልኤፍዎችን ከተለያዩ መለያዎች ጋር ይመሰርታል፣ እነዚህም በአንድ ላይ ከኤስ-አርናሴ በስተቀር ሁሉንም S-RNases ለይቶ ማወቅ ይችላል።9.
ምንም እንኳን አብዛኛዎቹ የዲፕሎይድ ድንች መስመሮች ከራሳቸው ጋር የማይጣጣሙ (SI) ቢሆኑም ከራሳቸው ጋር የሚጣጣሙ የዲፕሎይድ ድንች መስመሮች አሉ እና በዲፕሎይድ ድንች እርባታ መርሃ ግብሮች ውስጥ እራሳቸውን ተኳሃኝነትን ለማስተዋወቅ ሊያገለግሉ ይችላሉ ።10-12. ሆሳካ እና ሃኔማን አውራ S-locus inhibitor ካርታ ሰሩ (ስሊ) ጂን ከ ሀ Solanum chacoense በክሮሞሶም 12 የሩቅ ጫፍ ላይ መድረስ እና ድንች የተከተቡ መስመሮችን ለመፍጠር ተጠቅሞበታል13'14. በውጤታቸው መሰረት ሆሳካ እና ሃኔማን ይህን ሀሳብ አቅርበዋል ሲሊ ስፖሮፊቲክ እርምጃ ያለው እና ለዚያ ግብረ-ሰዶማዊነት ያለው የአበባ ዱቄት የተገለጸ ጂን ነው። ሲሊ ከግብረ-ሰዶማዊነት ጀምሮ ገዳይ ነው ስሊስሊ የጂኖታይፕስ ዓይነቶች በ F8 የኤስ. chacoense. ከእነዚህ ኤስ. chacoense (DS) - በራስ ተኳሃኝነትን ወደ ኤስ ለማስተዋወቅ የተፈጠሩ የተዳቀሉ መስመሮች። ቱቦሮሰም ዳራዎች. እዚህ, በዲፕሎይድ ድንች ውስጥ የራስ-ተኳሃኝነት ባዮሎጂን በተመለከተ ተጨማሪ ግንዛቤን ለማግኘት የእራስን ተኳሃኝነት መንስኤ ጂን መለየትን እንገልፃለን.
ውጤቶች እና ውይይት
በአንድ F2 ህዝብ መካከል ካለው መስቀል የተገኘ ሲሊ ለጋሽ (የተሰየመ ዲኤስ) እና ዳይፕሎይድ ኤስ. ቱቦሮሰም (D2) በክሮሞሶም 2 ላይ ለራስ-ቤሪ ስብስብ QTL መጠነኛ የሆነ ውጤት አስተውለናል፣ ነገር ግን ተከታዩ የድጋሚ ማጣሪያ የተሳካ አልነበረም። በርካታ የኤፍ 2 ህዝቦች በክሮሞሶም 12 ረጅም ክንድ አካባቢ ከፍተኛ የሆነ ውዥንብር እንዳሳዩ አስተውለናል፣ ግብረ-ሰዶማዊነት ለ DS-ያልሆኑ ሃፕሎታይፕ ሙሉ በሙሉ የለም።
በዚህ skewness እና ካርታ ላይ የተመሠረተ ሲሊ በክሮሞዞም 12 ላይ፣ ያንን መላምት አድርገናል። ሲሊ ጋሜቶፊቲካል በሆነ መልኩ ይገለጻል፣ ይህም ማለት እራስን በማዳቀል የእፅዋት heterozygous ለ ማለት ነው። ሲሊ (ስሊ/sli)፣ የበላይ የሆነውን የአበባ ዱቄት ብቻ የያዘ ሲሊ allele በራስ ማዳበሪያ ውስጥ መሳተፍ ይችላል። ይህንን መላምት ለመፈተሽ፣ ኃይለኛ እና ከፍተኛ ራስን ለም መስመር (16HP1-66) ወደ ኃይለኛ ራስን የማይስማማ መስመር (D16) ተሻግረናል፣ ሙሉ-ጂኖም ሁለቱንም በቅደም ተከተል አቅርበን እና የተገኘውን የF1 ህዝብ (17SC11) ተንትነናል። n = 251 ፣ ምስል 1ሀ, ለ) ይህ F1 ህዝብ ለብዙ ሎሲዎች ከፍተኛ ፖሊሞፈርፊክ በመሆኑ፣ ከመራባት ጋር የተያያዙትን ጨምሮ ብዙ አይነት ፍኖታይፕዎችን ተመልክተናል። ስለዚህ፣ የተኳሃኝነትን ፍኖተ-ዓይነት ግራ የሚያጋቡ የፅንስ መፈጠር ጉዳዮችን ለማስወገድ ከሁለቱም መስቀል እና ራስን ማዳቀል የተገኙ የቤሪ እና የዘር ፍሬዎችን እንዲሁም የአበባ ዱቄት ቱቦዎችን በሥዕሎች ውስጥ በማሳየት የሚያካትት በጣም ጥብቅ እና ጥብቅ የፍኖታይፕ ፕሮቶኮልን ተግባራዊ አድርገናል። ከአንድ በላይ የራስ-ቤሪን የሚያዘጋጁ ተክሎች እንደ ኤስ.ሲ ይቆጠራሉ, ነገር ግን ቢያንስ 10 እራስን የአበባ ዘር ካደረጉ በኋላ እራሳቸውን የቤሪ ፍሬዎችን የማያዘጋጁ ተክሎች, የእራስ የአበባ ቧንቧ እድገትን በአጻጻፍ ስልቱ ያሳያሉ, እና የአበባ ዱቄት ከተበከሉ በኋላ የመስቀል ፍሬዎችን ያስቀምጣሉ. የአበባ ዱቄት እንደ SI ይቆጠራሉ. በውጤቱም፣ የተኳኋኝነት ፍኖታይፕን ለማያሻማ ሁኔታ ለመገምገም የሚያስፈልጉት መስፈርቶች ስላልተሟሉ ጉልህ የሆነ የህዝቡ ክፍል ከጄኔቲክ ትንታኔዎች ተገለለ። አሁንም፣ የአብዛኛው ህዝብ 17SC11 ዘሮች የተኳሃኝነት ሁኔታ ሊገመገም ይችል እና ለራስ-ተኳሃኝነት (ተጨማሪ መረጃ 1) ለመለየት ታይቷል። ራስን ተኳሃኝነት ከ16HP1-66 የመነጨ በመሆኑ፣ የ KASP ማርከሮችን በክሮሞሶም 2 እና 12 ላይ የሚያነጣጥሩ SNPዎችን ለመንደፍ በ16HP1-66 heterozygous ነገር ግን በD16 ውስጥ ግብረ-ሰዶማዊነት የዚን ጂኖም ቅደም ተከተሎችን ለመንደፍ ተጠቀምን። ሲሊ በእናቶች ሚዮሲስ ውስጥ. የጄኔቲክ ካርታ ገንብተናል፣ የQTL ትንታኔን ሰርተናል፣ እና በክሮሞሶም 75.72 ረጅም ክንድ ላይ በጣም ጠቃሚ የሆነ QTL (LOD = 12) አግኝተናል (ምስል 1ለ) የሆሳካ እና የሃኔማን ውጤቶችን ማረጋገጥ.
QTLን በተለየ የዘረመል ዳራ ለማረጋገጥ፣ ከሶሊንታ የመራቢያ ፕሮግራም ከSI genotype D14 የተገኘ ሌላ በጣም ራስን ለም የሆነ ጂኖታይፕ ተሻግረን የተገኘውን የF1 ህዝብ (17SC25፣ ተጨማሪ መረጃ 1) ተንትነናል። ከ32SC17 የህዝብ ብዛት 25 ግለሰቦች መካከል ምንም SI ግለሰቦች አላገኘንም። የተለያየ ህዝብ ለማፍራት በጣም ለም የሆነውን ጂኖታይፕ መርጠን ወደ ሁለት የSI genotypes ተሻግረን በ17SC11 የህዝብ ብዛት ለይተናል፣ይህም የህዝብ ብዛት 18SC11 እና 18SC12 (ተጨማሪ መረጃ 1 እና ተጨማሪ ምስል 1)። እንደተጠበቀው፣ የሕዝብ 18SC11 እና 18SC12 ትንታኔ እንደሚያሳየው ሁለቱም ህዝቦች ለራስ ተኳሃኝነት ይለያሉ። እናቱን (17SC25-8) ለሙሉ ጂኖም ቅደም ተከተል አስገብተናል እና ይህንን መረጃ ተጠቅመን አዲስ የ KASP ማርከርን ለመንደፍ ለ17SC11 ህዝብ ጥቅም ላይ የዋለውን ተመሳሳይ አቀራረብ ተጠቅመን ነበር ፣ ግን በዚህ ጊዜ ክሮሞዞም 12 ን ብቻ ያነጣጠረ ነው። ቀጣይ የQTL ትንታኔ በሕዝብ ውስጥ ያገኘነውን QTL አረጋግጧል። 17SC11 ከ LOD እሴቶች 33.14 እና 120.94 በህዝብ 18SC11 እና 18SC12 በቅደም ተከተል (ተጨማሪ ምስል 2)።
ለመወሰን ፡፡ ሲሊ በጋሜቶፊቲካል የተገለጸ ነው፣ የF2 ህዝብን ተንትነናል (19SC1፣ n = 160) ለም እና ብርቱ ከሆነ 17SC11 ግለሰብ የተገኘ። በዚህ ህዝብ ውስጥ የአበባ እና የመራባት መጠን ከ F1 ጋር ሲነጻጸር ቀንሷል. በፍኖቲፒክ ትንተና ከ160 እፅዋት 81 እፅዋቶች እራሳቸውን የሚስማሙ ናቸው፣ 78ቱ ያልተወሰኑ (ND) ተብለው ተከፋፍለዋል በደካማ አበባ ወይም ደካማ የመራባት እና አንድ ተክል እራሱን የማይስማማ ውጤት አግኝቷል (ተጨማሪ መረጃ 1)። እኛ በወላጆች 12HP16-1 እና D66 ውስጥ ለተለዋጭ አለርጂዎች ግብረ-ሰዶማዊ የሆኑትን ኤስኤንፒዎችን በክሮሞዞም 16 ላይ የሚያነጣጥሩ የ KASP ማርከሮችን አዘጋጅተናል። ከጠቅላላው ክሮሞሶም ጋር 12 መለያየት ሬሾዎች ከሚጠበቀው 1፡2፡1 መለያየት በእጅጉ ይለያያሉ። በተጨማሪም፣ በራስ ተኳሃኝነት QTL ዙሪያ፣ ለወላጅ D16 ሃፕሎታይፕ ምንም ዓይነት ሎሲ ግብረ-ሰዶማዊነት የለም (ተጨማሪ ምስል 3)፣ በምትኩ 1፡1 መለያየት ለ heterozygous D16/16HP1-66፡ ግብረ-ሰዶማዊ 16HP1-66፣ ይህም መወገድ እንዳለበት የሚጠቁም ነው። የአበባ ዱቄት እጥረት ሲሊ የመለያየት መዛባትን ያስከትላል። ይህ የበላይነቱን የሚይዘው የአበባ ዱቄት ብቻ ነው የሚለውን መላ ምት ይደግፋል ሲሊ allele ራስን በማዳቀል ውስጥ ይሳተፋል። በተጨማሪም፣ ከአንድ ግለሰብ ውጪ፣ በSI እና SC መካከል ላለው ንፅፅር ፍኖታይፕ መደምደሚያ መደምደሚያ ነበር፣ ይህም ጥቅም ላይ የዋለው የፍኖታይፕ ፕሮቶኮል ጠንካራ እና ከስህተት የጸዳ መሆኑን ያሳያል።
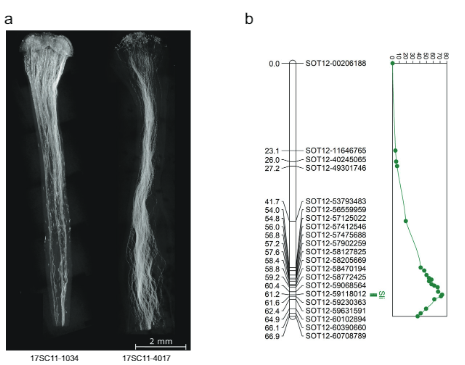
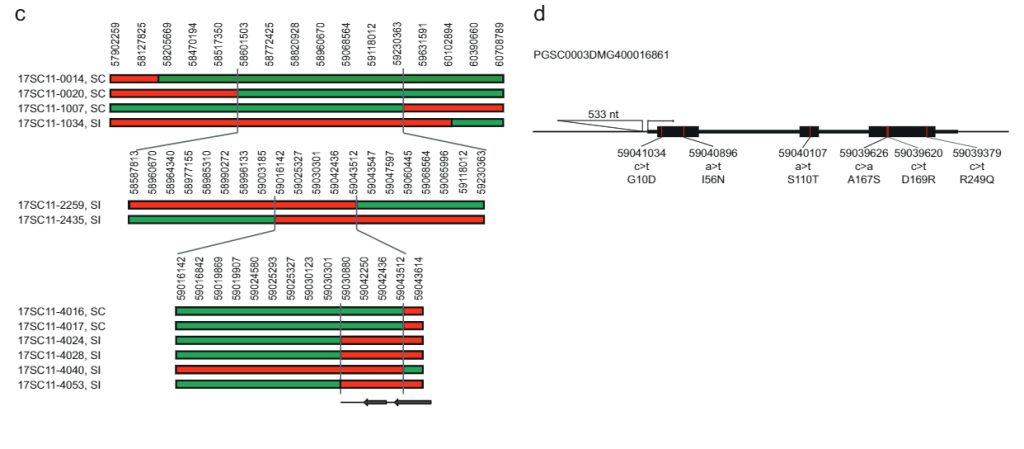
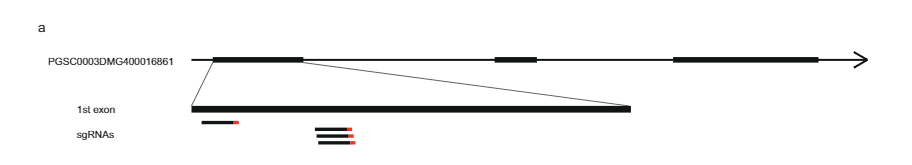
ሳለ 628 ኪ.ባ SLI በክሮሞሶም 12 ላይ ስሊ-አሌልን ከሕዝብ 17SC11 ተሸክሞ ወደ 169 ኪባ ወደ ትንሽ ተደራራቢ ክፍተት ቀንሷል በሕዝብ 18SC12፣ እነዚህ ክፍተቶች አሁንም ስሊ-ጂንን ለመለየት በጣም ትልቅ ነበሩ። ስለዚህ፣ የ S/i-ያካተተውን ክፍተት በእንደገና የማጣራት ዘዴ ለመቀነስ አላማን ነበር። በ ውስጥ ድጋሚ ውህደት ያላቸውን ተክሎች ለመለየት ሲሊ የጊዜ ክፍተት፣ 1374 17SC11 ችግኞችን በሁለት የ KASP ማርከሮች በቅርበት ድንበር እና ሁለት በሩቅ ድንበር ላይ አድርገናል። በሁለቱ የውጪ ጠቋሚዎች መካከል 81 ችግኞችን እንደገና በማጣመር ለቀጣይ የካርታ ስራ መርጠናል ። የማያሻማ ፍኖታይፕ ለማግኘት፣ እነዚህን ጂኖታይፕስ በእፅዋት በማባዛት በእያንዳንዱ ጂኖታይፕ ቢያንስ ሁለት ክሎኖች ላይ ፍኖተ-ታይፕ አደረግን። 81 ን ድጋሚ ውህዶችን በክፍተቱ ውስጥ ተጨማሪ ማርከሮችን በጂኖታይፕ ገለፅናቸው እና ሁለት መረጃ ሰጪ ድጋሚ ጥንብሮችን ለይተን ለይተናል እና ክፍተቱን ወደ 27.37 ኪባ የቀነሱ አምስት የተብራራ ጂኖች (ተጨማሪ መረጃ 1)።
ክፍተቱን የበለጠ ለመቀነስ ተጨማሪ 10165 ችግኞችን ከተመሳሳይ ህዝብ በአራት ጠቋሚዎች በዚህ 27.37 ኪ.ባ ርቀት ላይ በማጣራት 12 ሪኮምቢንቶችን ለይተናል። እነዚህ ተጨማሪ ጂኖታይፕ በ14 ተጨማሪ ማርከሮች ተቀርፀዋል እና ግልጽ የተኳኋኝነት ፍኖተ-ዓይነቶችን የሚያሳዩ ስድስት መረጃ ሰጪ ድጋሚ አካላትን ለይተናል። ሁለት ድጋሚ የ SC phenotype እና አንድ SI phenotype ያለው የ 27.37 KB ክፍተት የሩቅ ወሰን አረጋግጠዋል፣ ሶስት ዳግመኛ ከSI phenotype ጋር አዲስ የፕሮክሲማል ድንበር ሲወስኑ ክፍተቱን ወደ 12.6 ኪባ ብቻ የሚቀንስ ሁለት ጂኖች፣ PGSC0003DMG400016861PGSC0003DMG400016860 ምስል 1ሐ).
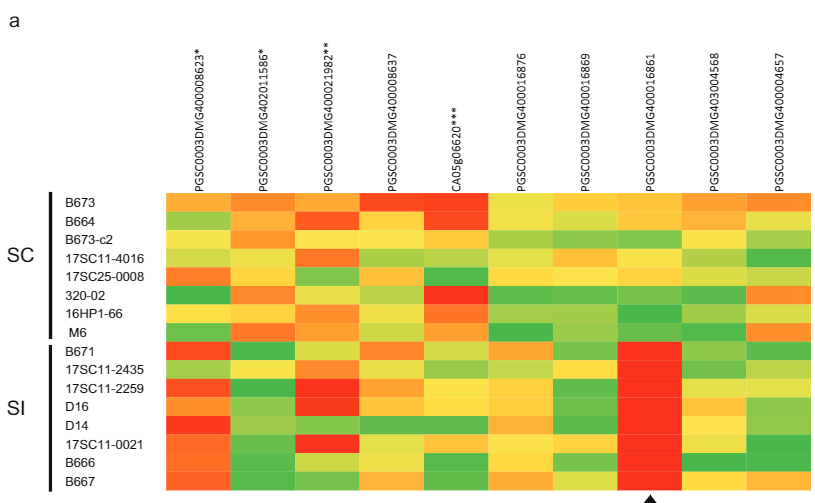
ለራስ-ተኳሃኝ ፍኖታይፕ ተጠያቂ የሆነውን እጩ ጂን ለመለየት የእነዚህን ሁለት ጂኖች ተከታታይ ልዩነት በበርካታ የሙሉ ጂኖም ተከታታይ ዳይፕሎይድ ድንች መስመሮች (ተጨማሪ መረጃ 2) ላይ ተንትነናል። ይህንን የቅደም ተከተል ልዩነት ከነዚህ መስመሮች SC/SI ፍኖተአይፕ ጋር በማነፃፀር ሁሉንም SC-ተኮር SNPs እና INDELS (ተጨማሪ መረጃ 2) ለይተናል። በመቀጠል፣ ሁሉንም ተመሳሳይ ያልሆኑ SNPዎችን በእጃችን ለይተናል እና የአሚኖ አሲድ መተኪያዎች በ Solanaceae ውስጥ ላሉ ተመሳሳይ ፕሮቲኖች የተለመዱ ወይም ልዩ መሆናቸውን ወስነናል። የእጩ ጂን PGSC0003DMG400016861 ስድስት SC-ተኮር የአሚኖ አሲድ ተተኪዎችን ያሳያል እና በተለይም የ 533 bp መግቢያ በ —108 bp ከመጀመሪያው ኮድን ላይ የሚገኝ ሲሆን ይህም SC allele ከSI allele ጋር ሲነጻጸር አገላለፅን እንደቀየረ ይጠቁማል። በእነዚህ የዘረመል ጥናቶች መሰረት PGSC0003DMG400016861 ነው ብለን ገምተናል። ሲሊ ጂን
ስሊ በአበባ ብናኝ ውስጥ መገለጹን ለማረጋገጥ ከ10 SI እና 10 SC የድንች ጂኖታይፕስ በብልቃጥ ውስጥ የአበባ ብናኝ አበቅልን እና አር ኤን ኤን ለ RNA ቅደም ተከተል አውጥተናል። ከቀሩት ሁለት እጩ ጂኖች፣ እጩ ጂን PGSC0003DMG400016861 ብቻ ነው የተገለፀው፣ ነገር ግን ከኤስ.ሲ. 2ሀ) በተጨማሪም በእጽዋት ውስጥ heterozygous ለ putative እጩ Sli ጂን, Sli allele ብቻ ተገልጿል. የሚገርመው፣ በChromosome 12 ላይ ከስሊ ቦታ አጠገብ የሚገኙ ሌሎች የአበባ ብናኝ የተገለጹ ጂኖች በ SC እና SI ተክሎች ውስጥ ተመሳሳይ የመገለጫ ደረጃዎችን አሳይተዋል (ምስል. 2ሀ) ስለዚህ, የ PGSC0003DMG400016861 ጂን ብቻ በተለይ በኤስ.ሲ. ተክሎች የአበባ ዱቄት ውስጥ ይገለጻል ብለን ደመደምን.
የ533 bp ማስገባትን አመጣጥ ለመመርመር፣ የ533 bp ቅደም ተከተል በNCBI ላይ የBLAST ፍለጋ አደረግን። የሚገርመው፣ ከSli-specific ingrediation (Sli-specific insertion) ጋር ተመሳሳይ የሆኑ ቅደም ተከተሎች በተከታታይ ኤስ. ቱቦሮሰም መለዋወጫዎች (ምስል. 2ለ) በተጨማሪም፣ 533 ቢፒ ማስገቢያ በኤስ. ፔኔሊሊ. ቅደም ተከተል ከኤስ. ፔኔሊሊ እንደ BLAST ጥያቄ በኤስ ውስጥ ተመሳሳይ ቅደም ተከተሎችን አግኝተናል። ሊኮፐርሲኩም. በ ውስጥ ያሉ ቅደም ተከተሎች phylogenetic ትንተና ኤስ, ኤስ. ፔኔሊሊ ና ኤስ. ሊኮፐርሲኩም ቡድኖች ኤስ. ፔኔሊሊ ቅደም ተከተል ከ ጋር ኤስ. ሊኮፐርሲኩም እና አንድ ኤስ ቅደም ተከተል፣ እነዚህ የጋራ መነሻን እንደሚጋሩ የሚጠቁም (ተጨማሪ ምስል 4 ሀ)። ማስገባቱ ከተንቀሳቃሽ አካል (TE) የተገኘ ነው ብለን ገምተናል። የነጥብ-ፕላት ግራፍ ከ 533 ቢፒሲ ቅደም ተከተል ፈጠርን እና ቅደም ተከተላቸው ጥቃቅን የተገለበጠ ድግግሞሾችን እንደያዘ ተመልክተናል (ተጨማሪ ምስል 4 ለ)። የ533 ቢፒኤስ ማስገባትን ከፋብሪካ MITE ዳታቤዝ አንጻር ለBLAST አስገብተናል፣ይህም ምክንያት ከ MITE ቤተሰብ DTA_Sot42 in ኤስ15, የሚያመለክተው 533 bp በፕሮፌሽናል ውስጥ ማስገባት ሲሊ በእርግጥ የመጣው ከቲኢ (ተጨማሪ ምስል 4 ሐ) ነው።
PGSC0003DMG400016861 በእርግጥ መሆኑን የበለጠ ለማረጋገጥ ሲሊእኛ የ SC allele exons የያዘ አገላለጽ ገንቢ ነድፈናል። ሲሊ በትውልድ አራማጁ እና ተርሚናተሩ መካከል (ምስል. 3ሀ) በቬክተር pBINPLUS (pBINPLUS-Sli)። ይህንን ግንባታ የተጠቀምነው ሁለት የSI genotypes ከካርታ ስራ 18SC12 ህዝብ ለመቀየር ነው። ከSI genotype B666 ከተገኙ ከእያንዳንዳቸው ከአምስቱ ነጻ ትራንስጀኒኮች ከሁለት እስከ ስድስት ክሎኖችን ፈጠርን እና ከSI genotype B667 የተገኙ ሶስት ትራንስጀኒክስ።
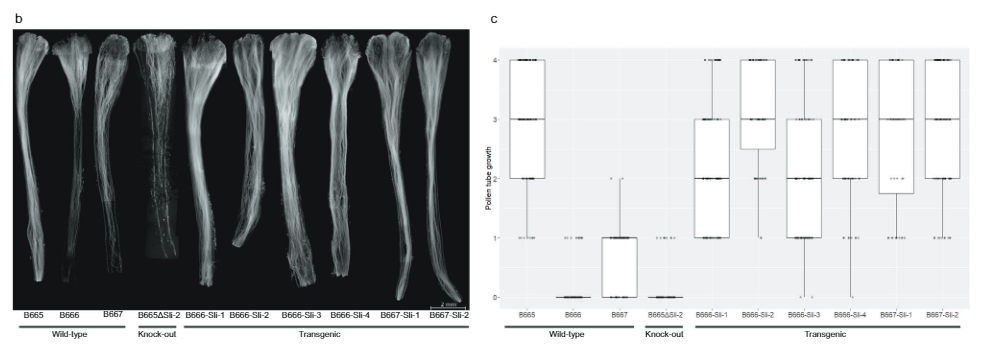
ከስድስት ገለልተኛ ትራንስጀኒክስ የመጡ ክሎኖች እራሳቸውን በሚበክሉበት ጊዜ ቤሪዎችን ያዘጋጃሉ (ተጨማሪ መረጃ 3)። በተጨማሪም, የፍሎረሰንት ማይክሮስኮፕ የአበባ ዱቄት ከ ሲሊ ትራንስጀኒክ እፅዋት ካልተቀየሩ ቁጥጥሮች ይልቅ ወደ ራሳቸው ዘይቤዎች ጠልቀው ያድጋሉ (ምስል. 3ለ፣ ሐ) በ 442 ራስን የአበባ ዱቄት ውስጥ የአበባ ዱቄት ቱቦ እድገት አስመዝግበናል። ሲሊ ትራንስጀኒክስ እና 179 እራስ-አበቦች በ 0-4 ሚዛን ባልተለወጡ መቆጣጠሪያዎች. አብዛኛዎቹ የአበባ ዱቄት ቱቦዎች በአብዛኛዎቹ ኦቭየርስ ላይ ደርሰዋል ሲሊ ትራንስጀኒክስ፣ ከመቆጣጠሪያዎቹ በጣም ትንሽ ክፍልፋይ ጋር ሲነጻጸር፣ PGSC0003DMG400016861 መሆኑን ያሳያል። ሲሊ ጂን
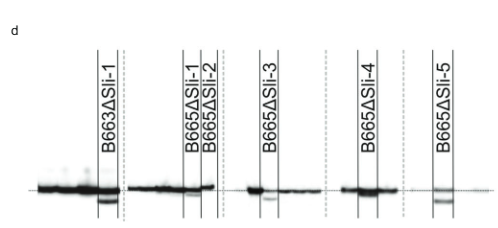
በመቀጠል፣ የ PGSC9DMG0003 (pAGM:CRISPRASli፣ Fig.400016861) ላይ ያነጣጠረ አራት የጂአርኤንኤን ኮድ የሚያሳይ የCRISPR-CasXNUMX ግንባታ ነድፈናል። 3ሀ) በዚህ ግንባታ ሁለት SC genotypes (B665 & B663) ቀይረናል እና 149 የተለወጡ ዳግም ማመንጫዎችን አግኝተናል። ከዚያ በCRISPR-Cas9 የተፈጠሩ INDELዎችን ለመለየት PAGEን በመጠቀም የታለመውን ኤክሶን ተንትነናል። እንደ አለመታደል ሆኖ፣ pAGM:CRISPRASli ቬክተር ዝቅተኛ ቅልጥፍና ነበረው፣ከ149 ተሃድሶዎች ውስጥ ስድስቱ ብቻ INDELs አሳይተዋል። ሲሊ (Fig. 3መ) ከእነዚህ CRISPR-Cas9 አምስቱ መስመሮች ለ INDELs heterozygous ናቸው፣ ግን አንዱ መስመር B665ASli-2 ለትንሽ INDEL ግብረ-ሰዶማዊ ነው። ያልተለወጠ B665 እራሱን የቤሪ ፍሬዎችን ያዘጋጃል እና ጥሩ የራስ-pollen ቱቦ እድገትን በ105 የተስተዋሉ ዘይቤዎች ሲያሳይ B665ASli-2 እራሱን በሚበከልበት ጊዜ ቤሪ አያዘጋጅም እና የአበባ ዱቄት በ 78 የተስተዋሉ ዘይቤዎች ማደግ አልቻለም (ምስል 3ለ፣ ሐ እና ተጨማሪ መረጃ 3)፣ PGSC0003DMG400016861 መሆኑን ተጨማሪ ማስረጃ በማቅረብ ሲሊ ጂን
በS-RNase ላይ በተመሰረቱ ጋሜቶፊቲክ ራስን አለመጣጣም ሲስተሞች ራስን ማዳበሪያ በፒስቲል ኤስ አር ኤን ኤ ወደ የአበባ ብናኝ ቱቦዎች ውስጥ ገብተው በራሳቸው የአበባ ዱቄት ወይም ተዛማጅ ኤስ-ሎከስ ኤፍ-ሣጥን (ኤስኤልኤፍ) በሌሉት የሳይቶቶክሲካል ተጽእኖዎች በፒስቲል ገለጻ ይከላከላል። ) ፕሮቲን. ተሻጋሪ ማዳበሪያ የነቃው በራሳቸው ያልሆኑ S-RNasesን ሊያውቁ እና ሊበክሉ በሚችሉ የአበባ ብናኝ ኤስኤልኤፍ ፕሮቲኖች ነው። እያንዳንዱ ኤስ-አሌሌ በርካታ SLF ዎችን ይመሰርታል ይህም እያንዳንዳቸው የተለየ S-RNaseን የሚያውቁ ሲሆን በአንድ ላይ በድንች ውስጥ የሚገኙትን አብዛኛዎቹን S-RNases ለይተው ማወቅ ይችላሉ S-RNase ከተመሳሳዩ S-allele በስተቀር። ስሊ የF-box ፕሮቲን PP2-B10ን ኮድ ያስቀምጣል፣ እሱም ከሌክቲን ጎራ ጋር የተገናኘ የF-box ጎራ። የሌክቲን ጎራዎች ከካርቦሃይድሬትስ ጋር መስተጋብር እንደሚፈጥሩ ይታወቃሉ፣ እና ከግላይኮሲላይድ ፕሮቲኖች ጋር መስተጋብር መፍጠር ይችሉ ይሆናል።16. በተጨማሪም, S-RNases glycosylated ታይቷል17. የ 533 bp ማስገባት በ SC allele of Sli አራማጅ ውስጥ የአበባ ብናኝ መግለጫን እንደሚያስችል እንገምታለን። ላይምቤር በድንች ውስጥ ስላለው የ MITE እንቅስቃሴ ዝርዝር ምርመራ 2% የሚሆኑት ከጂኒክ ክልሎች አቅራቢያ ከሚገቡት የ MITE መግባቶች በጂን አገላለጽ ላይ ከተደረጉ ለውጦች ጋር የተቆራኙ መሆናቸውን አረጋግጧል።
በተጨማሪም በ1935 ከተፈተኑት የባርኔጣ ህትመቶች ወደ ጄኒክ ክልሎች 13 ቱ የተዛመደውን ዘረ-መል (ጅን) ማስተካከል አስከትለዋል፣ ይህም የአበባ ዱቄትን ልዩ አገላለጽ መቀየሩን ያሳያል። ሲሊ በእውነቱ በ 533 bp በአስተዋዋቂው ውስጥ በመግባቱ ምክንያት ሊከሰት ይችላል።18. ቢሆንም, የዚህን መላምት ትክክለኛነት ለመወሰን ተጨማሪ ምርምር ያስፈልጋል.
ከዚህ ቀደም ክሎት እና ሌሎች በ 333 ኪ.ባ ልዩነት በክሮሞዞም 12 ለመለየት የጅምላ ሴግሬጋንት K-mer የካርታ አቀራረብን ተጠቅመዋል ስሊ የሚገኝበት19. እዚህ፣ የSli locusን በኤፍ 12 ህዝብ ውስጥ ወደሚገኝ ተመሳሳይ የክሮሞዞም 1 ክልል ካርታ አዘጋጅተናል እና ክፍተቱን ወደ 12.6 ኪባ ወደ 2 ጂኖች ለማሳነስ የድጋሚ ማጣሪያ ተጠቀምን። የአገላለጽ ትንተና እንደሚያሳየው ከእነዚህ ጂኖች ውስጥ የአንዱ SC allele በተለይ ከ SC genotypes ውስጥ በአበባ ዱቄት ውስጥ ተገልጿል. በመጨረሻም፣ ትራንስጀኒክ አገላለፅን በመጠቀም እና በCRISPR-Cas9-induced knock-out፣ PGSC0003DMG400016861 Sli መሆኑን በፍፁም እናሳያለን። የላይምቤር ጥናቱ እንደሚያሳየው ኤምአይቲዎች በቲሹ ተኮር በሆነ መልኩ ተቀራራቢ ጂኖችን መቆጣጠር እንደሚችሉ፣ በ Sli አስተዋዋቂው ውስጥ ያለው MITE መኖሩ የአበባ ብናኝ አገላለጽ መንስኤ መሆኑን ለማረጋገጥ ተጨማሪ ምርምር ያስፈልጋል።
በዚህ ጥናት ውስጥ በተመረመሩት ቁሳቁሶች (ተጨማሪ መረጃ 4) ፣ ከግብረ-ሰዶማዊነት ለ Sli ጋር የተዛመዱ ገዳይነት ሪፖርቶችን ቀደም ብለው ማረጋገጥ አልቻልንም ፣ ምክንያቱም አዋጭ የሆኑ F2 ተክሎች ለ Sli ቤሪዎችን ማዘጋጀት የቻሉ (ተጨማሪ መረጃ 1)12>20. በተጨማሪም ፣የኢንብሬድ-ላይን ሶሊንተስ ጂኖም ቅደም ተከተሎች ፣እንዲሁም ኢንብሬድ-መስመር M6 ፣ሁለቱም መስመሮች ለስላ ግብረ-ሰዶማዊ መሆናቸውን ያሳያሉ ፣ይህም ለስላይድ ግብረ-ሰዶማዊነት ገዳይ አይደለም ፣ምንም እንኳን አሁንም ገዳይ ሊሆን ይችላል። በቅድመ አያት ውስጥ ከስሊ ጋር በዘረመል የተገናኘ allele በእነዚህ ጂኖታይፕስ ውስጥ እንደገና በመዋሃድ ተወግዷል21'22. ነገር ግን፣ በዚህ ጥናት ውስጥ ከተገኘው መረጃ፣ በF2 ህዝብ ላይ የሚታየው የመለያየት መዛባት በክፍል ውስጥ ከኤስሊ ኤስአይኤሌል ጋር በተገናኘ ገዳይ አሌል ምክንያት ሊከሰት የሚችልበትን እድል ማስቀረት አንችልም። እስካሁን ድረስ Sli ራሱ S-RNasesን በቀጥታ ማወቅ እና መርዝ መቻል አለመቻል ግልጽ አይደለም። በተጨማሪም፣ Sli በሁሉም የ S-locus genotypes ውስጥ ራስን ተኳሃኝነት ያስገኝ እንደሆነ ግልጽ አይደለም። የአንዳንድ S-alleles ተግባር በ Sli ሊታገድ አይችልም. ይህንን ጥያቄ ለመፍታት ተጨማሪ ምርምር ያስፈልጋል.
የስላይን መለየት በዲፕሎይድ ድንች በመጠቀም የተዳቀለ መስመር ላይ የተመሰረተ ዝርያን ለማዳቀል ቢያስችልም፣ ሌሎች መሰናክሎች ግን ይቀራሉ። በመጀመሪያ ደረጃ, በጣም ጉልህ በሆነ መልኩ, ዳይፕሎይድ ድንች በዘር የሚተላለፍ የመንፈስ ጭንቀት ያጋጥመዋል, ይህም በመውለድ ላይ ጥንካሬን እና የመውለድ ችሎታን ይቀንሳል. በዲፕሎይድ የድንች መስመሮች ራስን ማዳበሪያ በቀጣይነት የሚያጠፉ አለርጂዎችን ማፅዳት የዘር ድብርትን ለመቀነስ ቀልጣፋ ዘዴ ሲሆን በአንፃራዊነት ጠንካራ እና ለም ድንች የተዳቀሉ መስመሮች እንዲፈጠሩ አድርጓል።2'23-25. ሁለተኛ፣ በዚህ ጥናት ውስጥ ጥቅም ላይ የዋለው የስሊ ለጋሽ DS፣ ከኤስ. chacoense ግንኙነት፣ ከኤስ የሚመጡ ጎጂ አሌሎችን በማገናኘት ወደ ችግሮች ሊያመራ ይችላል። chacoense. በ Solynta የዲፕሎይድ እርባታ ፕሮግራም ውስጥ ከኤስ ቻኮንሴስ በሚመጡ ጎጂ አሌሎች የተነሳ ግልጽ የሆኑ ጉዳዮችን አላየንም። በተጨማሪም፣ በራስ ተኳሃኝነት ላይ በቅርቡ የተደረገ ጥናት እንዳመለከተው፣ SC-specific k-mers ቀድሞውንም በበርካታ የ tetraploid cultivars ውስጥ ይገኛሉ፣ይህንን እምቅ ትስስር ጎትት ለማስወገድ የሚያስችል መንገድ እንደ Sli ለጋሾች ከነዚህ ዘሮች የሚመነጩትን ዲሃፕሎይድ በመጠቀም ነው።19.
ዘዴዎች
የእፅዋት ቁሳቁሶች. ሁሉም ያገለገሉ የእፅዋት ቁሳቁሶች በማሟያ መረጃ 4 ውስጥ ተዘርዝረዋል
የግሪን ሃውስ ሁኔታዎች. ሁሉም ተክሎች የሚበቅሉት ከ 14 ዲግሪ ሴንቲግሬድ በታች የሙቀት መጠኑ ሲቀንስ እና የሙቀት መጠኑ ከ 19 ዲግሪ ሴንቲግሬድ በላይ ሲጨምር መስኮቶችን በመክፈት በሚሞቁበት ጊዜ ነው. የብርሃን ጥንካሬ ከ 85 W/M2 በታች ሲወርድ ሰው ሰራሽ ብርሃን የተፈጥሮ ብርሃንን ያሟላል። ተክሎች የሚበቅሉት ከLentse Potgrond (Lentse Potgrond BV, Katwijk, ኔዘርላንድ) ልዩ በሆነ የድንች ንጣፍ ድብልቅ ውስጥ ነው. የሚፈለገውን የፒኤች መጠን ለማረጋገጥ የተመጣጠነ ውሃ ለመውሰድ፣ ለመሠረታዊ ቀስ በቀስ የሚለቀቅ ማዳበሪያ እና ኖራ ጥቅም ላይ የሚውለው የንዑስ ስትሬት ድብልቅ ከፔት ድብልቅ ነው። የንዑስ ውህዱ ማዳበሪያ 20፡20፡20 ናይትሮጅን፡ ፎስፈረስ፡ ፖታስየም መፍትሄን በመጠቀም ከኤሌክትሪካዊ ንክኪ (ኢ.ሲ.ሲ) 1.5 ነው።
ራስን ተኳሃኝነት ግምገማ. አበቦች እና ቡቃያዎች በሳምንት አንድ ጊዜ ይቆጠራሉ እና ጥንካሬ በወር አንድ ጊዜ ከ 1 እስከ 9 ባለው ሚዛን 1 በጣም ኃይለኛ ያልሆነ ተክል ሲሆን 9 በጣም ኃይለኛ ተክል ነው። ከአንድ ተክል የበርካታ አበባዎች የአበባ ዱቄት በ Eppendorf ቱቦ ውስጥ ተሰብስቦ ወዲያውኑ በተመሳሳይ አበባዎች ላይ እራስን ለማዳቀል በሳምንት ቢበዛ 10 አበባዎች ጥቅም ላይ ይውላል. በእያንዳንዱ የራስ-ቤሪ ቢያንስ 35 ዘርን የያዙ ከሁለት በላይ የራስ-ቤሪዎችን ያዘጋጁ ተክሎች ከራሳቸው ጋር ተኳሃኝ ተብለው ተመድበዋል። የሴቶችን የመራባት ችሎታ ለመወሰን ቢያንስ ከሶስት የማይዛመዱ የጂኖታይፕ ዝርያዎች ውስጥ ተክሎች በጅምላ የአበባ ዱቄት ተበክለዋል. ቢያንስ 10 እራስን የአበባ ዘር ካደረጉ በኋላ የራስ ፍሬዎችን ያላዘጋጁ፣ ነገር ግን ቢያንስ አንድ የጅምላ ቤሪ ያዘጋጁ እና ለም የአበባ ዱቄት በአጉሊ መነፅር በራሳቸው የተበከሉ ቅጦች ላይ ያሳዩ እፅዋት ከራሳቸው ጋር የማይጣጣሙ ተብለው ተመድበዋል። የቤሪ እና የዘሩ ስብስብ መረጃ የማያሳምም ከሆነ ከካርታው ህዝብ ለ 40 ጂኖታይፕስ (17SC11: n = 14፣ 18SC11፡ n = 7፣ 18SC12፡ n = 19)፣ ፍኖቲፒካል አመዳደብ በራስ የአበባ ዘር እድገት ላይ የተመሠረተ ነበር በቅጦች (በተጨማሪ መረጃ 1 ላይ የተጠቀሰው)።
የቅጥ ምስል. የአበባ ዱቄት ቱቦ እድገትን በዓይነ ሕሊና ለማየት, የአበባ ዱቄት ከተሰራ በኋላ ከ24-48 ሰአታት ውስጥ የተበከሉ ቅጦች ተወግደዋል ከዚያም በ 3: 1 ኢታኖል: አሴቲክ አሲድ ቢያንስ ለ 24 ሰዓታት ተስተካክለዋል. ከዚያም ስልቶቹ በ 8M NaOH ውስጥ ለ 10 ደቂቃዎች በ 65 ዲግሪ ሴንቲ ግሬድ ውስጥ ተጨፍጭፈዋል እና ሁለት ጊዜ በዲዮኒዝ ውሃ ታጥበዋል. ቅጦች በአጉሊ መነጽር ስላይዶች ላይ ተቀምጠዋል እና በ 2MK ውስጥ 5% Aniline blue (Carl Roth GmbH) በመጠቀም ለ0.1-0.1 ደቂቃ ቆሽሸዋል4P2O7 (pH = 7)፣ ከዚያም የሽፋን መሸፈኛ በመጠቀም በ glycerol ውስጥ ተጨፈጨፈ እና የማጣሪያ ስብስብ 01 (BP 365/12፣ FT 395 እና LP 397) በመጠቀም Zeiss Axiolab fluorescence ማይክሮስኮፕ በመጠቀም ተመልክቷል። ሁሉም ቅጦች የተስተዋሉ እና የተመዘገቡት ሁለት ግቤቶችን በመጠቀም ነው፡ (1) ወደ ስታይል ውስጥ ዘልቆ መግባት፣ ከፍተኛ መጠን ባለው መቶኛ እንደተገለጸው፣ (2) % የአበባ ዱቄት ቱቦዎች ወደ ጥልቅ ዘልቆ ደርሰዋል። ከዚያም እነዚህን መቶኛዎች ወደ 0-4 ሚዛን ቀይረናል፣ ምንም አይነት የአበባ ዱቄት ቱቦዎች እንቁላል ያልደረሱባቸው ቅጦች 0 ነጥብ አግኝተዋል፣ ከ0 እስከ 25 በመቶው የአበባ ዱቄት ቱቦዎች 1 ነጥብ ያገኙበት፣ ቅጦች በ25 መካከል እና 50% የአበባ ዱቄት ቱቦዎች እንቁላል ውስጥ ደርሰዋል 2 ነጥብ አግኝተዋል, ከ 50 እስከ 75% የአበባ ዱቄት ቱቦዎች እንቁላል ውስጥ የደረሱባቸው ቅጦች 3 ነጥብ አግኝተዋል, እና ከ 75% በላይ የአበባ ዱቄት ቱቦዎች ወደ እንቁላል ውስጥ የደረሱበት ቅጦች ነጥብ 4.
የምስል ማግኛ።. የተመረጡ ቅጦች የዚስ አክሲዮፎት ማይክሮስኮፕ ከማጣሪያ ስብስብ 01 ጋር፣ Zeiss AxioCam ICc 5ን በመጠቀም ተቀርፀዋል። ምስሎቹ የተቀረጹት የዚስ ዜን 2.3 (ሰማያዊ እትም) ሶፍትዌር ጥቅል በመጠቀም ነው። በግዢ ወቅት ቅንብሮቹ ዳራውን ለመቀነስ ተስተካክለዋል። ቅጦች የተቀረጹት የ x5 ዓላማን በመጠቀም ነው እና እንደ TIFF ፋይሎች የተቀመጡ ሲሆን በ 2464 x 2056 ፒክስል ጥራት ባለ 24-ቢት ጥልቀት። እስከ ስምንት የሚደርሱ የተለያዩ ምስሎች Panavue image assembler በመጠቀም ተሰብስበዋል። የበለስን ለመፍጠር የተገጣጠሙ ቅጦች ንፅፅር እና ብሩህነት ተስተካክሏል. 1ሀ እና 3c.
የዲኤንኤ ማውጣት. በአምራቹ የቀረበው ፕሮቶኮል መሰረት ለKASP የካርታ ህዝብ ትንተና፣ የቅጠል ናሙናዎች ወደ VHLGenetics (ዋገንገን፣ ኔዘርላንድ) ዲኤንኤ ለማውጣት sbeadex™ ኪት (LGC Genomics GmbH፣ Berlin፣ Germany) ተልከዋል።
የ KASP ትንተና. የኮምፕቲቲቭ አሌል-ተኮር PCR (KASP™) ትንታኔ በVHLGenetics (ዋገንገን፣ ኔዘርላንድስ) የተከናወነው በእኛ ቁስ ውስጥ ለሚለያዩ SNPs የተለየ እንዲሆን የተነደፉትን KASP assays በመጠቀም ነው። የ KASP ሙከራዎች የተካሄዱት በአምራቹ (LGC Genomics GmbH, በርሊን, ጀርመን) በቀረበው ፕሮቶኮል መሰረት ነው. ትክክለኛ መለያየትን እና የጂኖታይፕ ጥሪን ለማረጋገጥ ከKASP አሰሳ ውጤቶች SNPviewer (በ lgcgroup.com/products/genotyping-software/snpviewer ላይ ይገኛል) በመጠቀም ታይተዋል።
የግንኙነት ትንተና. ራሳቸውን የሚስማሙ የሴት ወላጆች ሃፕሎታይፕ በተለያዩ SNPs መካከል የመልሶ ማጣመር መጠኖችን በመተንተን ከጂኖታይፕ መረጃ እንደገና ተገንብተዋል። ይህ ውሂብ የ SNP ጥሪዎችን ወደ "axb" ቅርጸት ለመቀየር ጥቅም ላይ የዋለ ሲሆን በዚህ ውስጥ "a" ሃፕሎታይፕ ከራሱ ጋር ከሚስማማው የስሊ አሌል ጋር የተገናኘ ሲሆን የ"b" ሃፕሎታይፕ ግን ከራሱ ከማይስማማ የስሊ አሌል ጋር የተገናኘ ነው። የማገናኛ ካርታዎች የተፈጠሩት Joinmap 4.1ን በመጠቀም ነው።26 በሕዝብ ዓይነት DH እና በነባሪ ቅንጅቶች።
የ QTL ካርታ ስራ. የፍኖታይፕ ውሂቡ ወደ አሃዛዊ ባህሪ ተለውጧል 1 ለእያንዳንዱ ለራስ-ተኳሃኝ ጂኖታይፕ፣ 0 ለእያንዳንዱ ለራሱ የማይስማማ ጂኖታይፕ፣ እና * ወደ ጂኖታይፕ ተኳሃኝነት ሊታወቅ አልቻለም። የQTL ካርታ ስራ የተከናወነው በ MapQTL ውስጥ ያለውን የጊዜ ክፍተት ካርታ በመጠቀም ነው።27. የ MapQTL ውጤቶች የ QTL ቦታዎችን በ Mapchart 2.3 ለመፍጠር ጥቅም ላይ ውለው ነበር።28.
ባዮኢንፎርማቲክ ትንታኔ. በመጀመርያው 27.37 ኪ.ባ ልዩነት ውስጥ ትክክለኛ የጂን ሞዴሎችን ለመለየት ለዲኤም4.04 የማጣቀሻ ጂኖም፣ የPGSC ማብራሪያ እና የITAG ማብራሪያ ሁለት የተለያዩ የጂን ማብራሪያዎችን መርምረናል። ሂርሽ እና ሌሎችንም ይመልከቱ።29. የማብራሪያዎቹን ትክክለኛነት ለማረጋገጥ ከሁለቱም ማብራሪያዎች ከተገመቱት የፕሮቲን ቅደም ተከተሎች ጋር የ BLASTp ፍለጋዎችን አድርገናል። በBLASTp ፍለጋ ምርጦችን ከጥያቄያችን ጋር በማነፃፀር፣ በተተነበየው የፕሮቲን ቅደም ተከተል ውስጥ ያሉ ሁሉም የተብራሩ ኤክስፖኖች እና ጎራዎች በድንች እና በሌሎች የእፅዋት ዝርያዎች ውስጥ ባሉ ተመሳሳይ ፕሮቲኖች የተደገፉ መሆናቸውን ወስነናል። በተጨማሪም በSPUD DB ላይ በአደባባይ የሚገኙ አር ኤን ኤ-ሴክ ቤተ-መጻሕፍት (በsolanaceae.plantbiology.msu.edu/cgi-bin/ gbrowse/potato/) እና NCBI ጂኖም መረጃ መመልከቻ (በ ncbi.nlm.nih.gov/genome/gdv ላይ ይገኛል) /አሳሽ/) የተለጠፈ ኤክስፖኖች የገለጻ ማስረጃ እንዳላቸው ለመወሰን ጥቅም ላይ ውለዋል። እነዚህ ሁለት አቀራረቦች በሁለቱም ማብራሪያዎች ውስጥ የጂን ሞዴሎችን ኢንትሮን-ኤክስን አወቃቀሮችን እንድናረጋግጥ አስችሎናል፣ በዚህም ምክንያት አንድ ወይም ከዚያ በላይ የጂን ሞዴሎች በጥያቄ ውስጥ ያለውን ጂን ለመወከል በመረጃ ላይ የተመሰረተ ምርጫ አስገኝተናል። በእነዚህ አካሄዶች ላይ በመመስረት፣ እጩ ጂን PGSC0003DMG400016862 ከፊል እና ትርጉም በማይሰጥ መልኩ ተገለፀ እና ከተጨማሪ ትንታኔዎች ተወግዷል። የጂን ሞዴል Sotub12g029970 ትክክል ነው ተብሎ ሲታሰብ የPGSC አቻው PGSC0003DMG400016860 ተቆርጧል። በአብዛኛው ከተጠቀሰው የጊዜ ክፍተት ውጭ ስለሚገኝ እና በ SC እና SI ተክሎች መካከል ምንም ተዛማጅ የአሚኖ አሲድ መለወጫዎች ሊታወቁ አልቻሉም, ይህ ጂን ከተጨማሪ ትንታኔዎች ተወግዷል.
ተለዋዋጭ ትንተና. በ 27.37 ኪ.ባ ልዩነት ውስጥ ሚውቴሽን ለመለየት ለራስ-ተኳሃኝ ጂኖታይፕስ ሁሉም ከፍተኛ መተማመን SNPs (ተጨማሪ መረጃ 3) በ DS ፣ 1SC17-100 እና 18SC17-100 (ሦስቱም ግብረ ሰዶማውያን ስለሆኑ) ተወስኗል። የ SC allele የ ስሊ (ስሊ/ስሊ')')፣ (2) ግብረ-ሰዶማዊነት በD16 (ምክንያቱም D16 ለ SI allele of Sli (sli/sli)) እና (3) ሄትሮዚጎስ በሁለቱም 16HP1-66 እና 17SC25-8 (ሁለቱም ለ SC allele Sli (Sli) heterozygous ናቸውና። /sli))። የ Allelic ቅደም ተከተል የተገኘው በ SPAdes ስሪት 3.11.1 በመጠቀም በዲ ኖቮ ስብሰባ ነው።30 የ150 nt paired-end የኢሉሚና መረጃ ከላይ የተዘረዘሩት እፅዋት (በግምት 25-30X ተከታታይ ጥልቀት)። የውጤት ማመሳከሪያዎች ከዲኤም ማጣቀሻ (ሚኒማፕ2 ስሪት 2.1 በመጠቀም) እና በአስተማማኝ ሁኔታ ከ27 ኪባ ጋር ለሚጣጣሙ ተጣርተዋል። ከእነዚህ የተጣጣሙ ውዝግቦች፣ ከDM4.03 አንጻር ያለው ልዩነት በቁጥር ተቆጥሯል (የ subroutines mpileup እና ጥሪ ከbcftools፣ ስሪት 1.9) እና በተለዋዋጭ የጥሪ ቅርጸት (VCF) ተዘርዝሯል።
የአሚኖ አሲድ ለውጥ ትንተና. ከዚህ የ SC-specific ሚውቴሽን ዝርዝር ውስጥ ሁሉም ተመሳሳይ ያልሆኑ SNPs ከተሰየሙት የኮድ ኤክስፖኖች ጋር በመደራረብ ተለይተዋል። አሚኖ አሲድ ከዲኤም ወይም ከ SI ቅደም ተከተል አንጻራዊ ለውጦች ተዘርዝረዋል። የBLASTp ፍለጋዎችን የፕሮቲን ቅደም ተከተል በመጠቀም እና ከፍተኛ 100 BLASTp ስኬቶችን በመጠቀም በርካታ ተከታታይ አሰላለፍ በማከናወን ልዩ የአሚኖ አሲድ ለውጦች ተለይተዋል።
በአራማጅ እና ተርሚናል ክልሎች ውስጥ ልዩነት. ከፍተኛው 1500 nt ያለው የላይኛው ጂን ኮድ ኮድ ቅደም ተከተል እስከሚሰጠው ድረስ የማስተዋወቂያው ክልል የጀማሪ ኮዶን ወደ ላይ ያለው ቅደም ተከተል እንዲሆን ተመርጧል። በአስተዋዋቂ ክልሎች ውስጥ አስደናቂ ልዩነት በ27.37 ኪ.ባ ክፍተት ውስጥ ተገኝቷል፣ ከእነዚህም ውስጥ በጣም አስደናቂው በርካታ ትላልቅ ስረዛዎች እና ከአስር እስከ በመቶዎች የሚቆጠሩ ኑክሊዮታይድ ርዝማኔዎች መጨመር ናቸው። ከዲኤም ጋር በተዛመደ በ Sli ክፍተት ውስጥ ያሉ ሁሉም ልዩነቶች የተገኙት፣ የፕሮሞተር/ወደላይ ክልል እንዲሁም የተርሚናተር/የታችኛው ተፋሰስ ክልልን ጨምሮ።
የአበባ ዱቄት ማግኘት እና ማብቀል. በስእል ውስጥ ከተዘረዘሩት የጂኖታይፕ ዝርያዎች የአበባ ዱቄት. 2a በኤሌክትሮኒካዊ የጥርስ ብሩሽ በመጠቀም ክፍት አበባዎችን በማንቀጥቀጥ እና በ 1.5 ml Eppendorf tubes ውስጥ የአበባ ዱቄት በመሰብሰብ ተገኝቷል. ከግዢው በኋላ የአበባ ዱቄት ክፍት የሆኑትን የ Eppendorf ቱቦዎች የአበባ ዱቄት በአየር በታሸገ ሣጥን ውስጥ ሲሊካ ጄል በክፍል ሙቀት ውስጥ ለ 24 ሰዓታት በማከማቸት ደርቋል. ከዚያ በኋላ የአበባ ዱቄት በ -20 ዲግሪ ሴንቲ ግሬድ ውስጥ እስከ ተጨማሪ ጥቅም ላይ ይውላል.
የአበባ ዱቄት በ 2.5 ሚሊር ፈሳሽ መካከለኛ (5% (ወ/ቪ) sucrose, 9 mg/l Boric acid, 50 mg/l CaCly73.5H ውስጥ 2 ሚ.ግ የደረቀ የአበባ ዱቄት በማገድ ተበቅሏል.2ኦ፣ 118 mg/l Ca (NO3) 24 ህ2ኦ፣ 123 mg/l MgSO.4VH2ኦ) በ 3.5 ሴ.ሜ ዲያሜትር የፔትሪ ምግቦች በፓራፊልም የታሸጉ. የአበባ ዱቄት በፔትሪ ሳህኖች ውስጥ ለ 24 ሰዓታት በጨለማ ውስጥ በሚንቀጠቀጥ ኢንኩቤተር ውስጥ በክፍሉ የሙቀት መጠን እና በ 125 RPM ይንቀጠቀጣል. የበቀለውን የአበባ ዱቄት የያዘው ፈሳሽ መካከለኛ ወደ 2 ሚሊ ሜትር የኤፔንዶርፍ ቱቦዎች በጥንቃቄ የተገጠመ የፔፕቴት ምክሮችን በመጠቀም የአበባውን ቱቦዎች እንዳይበላሹ የመክፈቻውን መጠን ለመጨመር ተሻሽለዋል. የ Eppendorf ቱቦዎች በ 600xg ለ 1 ደቂቃ ሴንትሪፉድ ተደርገዋል እና መካከለኛው በቧንቧ በጥንቃቄ ተወግዷል. እንክብሉ እና አንዳንድ የቀሩት መካከለኛዎች ወዲያውኑ በፈሳሽ ናይትሮጅን ውስጥ በረዶ ተደርገዋል ፣ ሁለት አይዝጌ ብረት ዶቃዎች (2 ሚሜ ዲያሜትር) ተጨምረዋል እና ናሙናዎቹ በTissueLyser II (Qiagen GmbH ፣ Hilden ፣ Germany) በ 20 Hz ለ 1 ደቂቃ ተፈጭተዋል።
አር ኤን ኤ ማውጣት እና ቅደም ተከተል. Buffer RLT (Qiagen GmbH) ወደ የተፈጨ የአበባ ዱቄት ናሙናዎች ተጨምሯል እና ናሙናዎቹ በረዶ እንደቀሩ በማረጋገጥ። አር ኤን ኤ ማውጣት የተከናወነው በአምራቹ ፕሮቶኮል (Qiagen GmbH, Hilden, Germany) መሰረት RNeasy ሚኒ ኪት በመጠቀም ነው። የ250-300 ቢፒ ማስገቢያ መጠን ሲዲኤንኤ ቤተ-መጻሕፍት በቅደም ተከተል 150nt የተጣመሩ-መጨረሻ ንባብ፣በናሙና ከ30-42 ሚሊዮን ንባብ ጥንዶች (ኖቮጂን፣ ካምብሪጅ፣ ዩናይትድ ኪንግደም) ሰጡ።
ሌሎች የ RNA-seq የውሂብ ስብስቦች. የ(ቲሹ-ተኮር) አገላለጽ ደረጃዎች አጠቃላይ እይታ ለመፍጠር ሁሉም የተጣመሩ-መጨረሻ ተከታታይ አር ኤን ኤ-ሴክ ዳታ ስብስቦች እንደ ኦርጋኒዝም መለያ ተሰጥተዋቸዋል "Sola-num tuberosum" ከህዝብ ጎራ ወርደዋል (NCBI-SRA፣ ቀን 2018/17/13)፣ በድምሩ 441 የተጣመሩ fastq የውሂብ ስብስቦች። ከእነዚህ 441 የህዝብ መረጃ ስብስቦች 3 የተፈጠሩት ከስታይል ቲሹ (SRR7402817-SRR7402819) እና ሌሎች ሁሉም ከተለያዩ የአበባ ዱቄት ያልሆኑ ቲሹዎች፣ የእድገት ደረጃዎች እና የእፅዋት ውህዶች ነው።
የ Solyntus ማጣቀሻ ስብሰባ. ለአገላለጽ ትንተና፣ በቅርቡ የተገኘው የግብረ-ሰዶማውያን ማመሳከሪያ መስመር Solyntus (ስሪት 1.0፣ በ ላይ ሊወርድ ይችላል)። www.plantbreeding.wur.nl/Solyntus/) እንደ ማመሳከሪያ ጂኖም ጥቅም ላይ ውሏል. Solyntus እንደ ሶሊንታ የመራቢያ ፕሮግራም አካል ሆኖ የተፈጠረ በመሠረቱ ግብረ-ሰዶማዊ ዝርያ ነው።21. በዚህ ጥናት ውስጥ ያሉት የካርታ ክፍተቶች ከዲኤም v4.03 ጂኖም ስብሰባ የተገመቱ ናቸው።31 ወደ Solyntus 1.0 ጂኖም ስብሰባ በመሠረታዊ ተመሳሳይነት ፍለጋዎች (BLASTn እና bedtools በመጠቀም) በ (Solyntus 1.0 ጂኖም የመሰብሰቢያ መጋጠሚያዎች) 53532708-53954293 (Interval I, 421.6kb <—628.9kb, 53683239kb), 53867377terval 184.1-168.7 kb<- 53731620kb)፣ 53763003-31.4 (በመሃል III፣ 27.4kb<-53753977kb) እና 53763003-9.0 (Interval IV፣ 12.6kb <-1.0 kb) በቅደም ተከተል። በቅንፍ መካከል ያለው ተከታታይ የካርታ ክፍተት ቁጥር [Solyntus 1.0 መጋጠሚያዎች]፣ መጠን በ Solyntus-4.03 እና መጠን በዲኤም-4.04/4.03፣ በቅደም ተከተል። ሁሉም ክፍተቶች በክሮሞሶም ST12ch12_RaGOO (ክሮሞዞም 1.0 መሆን) ላይ ይገኛሉ እና በ Solyntus 4.03 ስብሰባ ውስጥ አንድ ክፍተት አልያዙም። የጊዜ ክፍተት ልዩነት በበርካታ ክፍተቶች (ኤን) በተዛመደ የዲኤም ቅደም ተከተል እና በሁለቱም ጂኖም መካከል ባለው ሰፊ ልዩነት ይከሰታል። በዲኤም ጂኖም (DM-4.04/12) ላይ ተጓዳኝ ክፍተቶች፡ የጊዜ ክፍተት I፡ chr58601503፡59230363-12፣ የጊዜ ክፍተት II፡ chr58962004፡59130723-59016142; ክፍተት III: 59043512-12; የጊዜ ክፍተት IV: chr59030880: 59043512-XNUMX.
በሶሊንተስ 1.0 ላይ ያለው የጂን ማብራሪያ ከሶስት የተለያዩ የጂን ካታሎጎች የተገመተ ነው (ድንች DM4.03፣ ITAG4.0 የቲማቲም ጂኖም ማብራሪያ የሴፕቴምበር 6፣ 2019 የተለቀቀው)32, እና Pepper-v. 1.5533GeMoMa (v1.6.1) ን በመጠቀም በሶሊተስ ጉባኤ ላይ ተቀርጿል። ይህ የተደረገው በግለሰብ የጂን ካታሎጎች ውስጥ ያሉ ጉድለቶችን ለማካካስ እና ሊኖሩ ስለሚችሉ ጂኖች እና/ወይም የተገለጹ ሎሲዎች ስለመኖራቸው ግንዛቤያችንን ከፍ ለማድረግ ነው።
RNA-seq የማንበብ ካርታ እና የጽሑፍ ብዛት ብዛትfiመጥቀስ. ሁሉም 5 SC፣ 3 SI እና ሁሉም 441 የህዝብ አር ኤን ኤ ሴክ ዳታ ስብስቦች hisat2 (ስሪት 2.1.0) በመጠቀም ወደ Solyntus ማጣቀሻ ጂኖም ተቀርፀዋል። GeMoMaን በመጠቀም የተገኘው ድቅል ዘረ-መል ካታሎግ በትራንስክሪፕት ለሚመራ የተትረፈረፈ ግምት StringTie (ስሪት 2.1.1) ከቅንጅቶች -t -c 5 -f 0.05 -G እና GeMoMa የተጠቃለለ Solyntus1.0 gff ፋይልን በመጠቀም ጥቅም ላይ ውሏል። ሁሉም የተስተዋሉ አገላለጾች በ 500 ኪ.ባ ክፍተት ዙሪያ ሲሊ ቦታ እንደ ማእከል ተገምግሟል፣ በጠቅላላው 90 (የተገመቱ) የጂን ሎሲዎች ይገኛሉ። በ SC ናሙናዎች ውስጥ ከነዚህ የዘረመል ቦታዎች ውጭ ምንም የሚታይ አገላለጽ አለመኖሩን አረጋግጠናል። በ 500 ኪ.ባ ክፍተት ውስጥ፣ ከላይ እንደተገለጸው ከካርታ ክፍላችን I-IV ጋር ስንገናኝ በመቀጠል አነስተኛውን የእጩ ጂኖች አመልክተናል።
በfiየ haplotype-speci እርማትfiሐ አገላለጽ. ከ90 የተገለፀው ሎሲ በ500 ኪባ ክፍተት፣ በሁሉም የ SC/SI ናሙናዎች ውስጥ ከተመረጠው ከ8 FPKM ገደብ በላይ 20 ብቻ ተገልጸዋል። እነዚህን ጣቢያዎች ሃፕሎታይፕ-ተኮር (Sli ወይም sli) የመግለፅ ደረጃ ልዩነቶችን ለመለካት ተጠቀምን። የተመረጠው የቃላት ገደብ በቂ የንባብ ጥልቀት ውሎ አድሮ እና በአስተማማኝ ሁኔታ አገላለፁን ወደ 2 ሃፕሎታይፕ ደረጃ ለማድረስ አስችሏል። ከፒኤስሲ ሎከስ ራሱ ጋር (በSI ተክሎች ውስጥ መግለጫ የሌላቸው)፣ እነዚህ 8 + 1 ሎሲዎች በእያንዳንዱ 8 ናሙናዎች (SAMtools phase ስሪት 1.7፣ ነባሪ መቼቶች) ሃፕሎታይፕ ተደርገዋል። የተገኙት ሃፕሎታይፕድ (የተጣመሩ) fastq ፋይሎች ዲ ኖቮ የተሰበሰቡት SPAdes (ስሪት 3.11.1) በመጠቀም ነው። የተገኙት ኮንቲግዎች ለብዙ ብዛት ተጣርተው ሙሉ ርዝመት ያላቸው ኤምአርኤንኤዎች ተደርገው ይገመታሉ፣ ይህም ከዋናው (ሃፕሎታይፕድ) የአይሶፎርም ጋር ይዛመዳል። በአንዳንድ ሁኔታዎች፣ ይህ በአማራጭ የተከፋፈሉ ኢሶፎርሞችን አስወገደ፣ አንዳቸውም በበቂ ንባብ የተደገፉ ምንም ግልጽ ባዮሎጂያዊ ጠቀሜታ የላቸውም። በእነዚህ ሃፕሎታይፕድ ኤምአርኤንኤ ቅደም ተከተሎች ያለው ልዩነት አንድ ወይም ሁለቱም ሃፕሎታይፕዎች በእያንዳንዱ ተጓዳኝ ሎሲ/ናሙናዎች ውስጥ መገለጣቸውን ለማረጋገጥ (ለማረጋገጥ) ጥቅም ላይ ውሏል።
ንድፍ ሲሊ አገላለጽ መገንባት. የስሊ አገላለጽ ካሴትን ለመንደፍ የSli donor plant DSን ቅደም ተከተል ተጠቅመንበታል። የአፍ መፍቻ መግለጫን ለመፍቀድ ፒኤስሲ፣ ቤተኛ አራማጁን (ከመጀመሪያው ኮድን 1563 ቢፒ ወደላይ)፣ ሦስቱ ኤክሰኖች እና ቤተኛ ተርሚነተር (740 ቢፒ ከስቶል ኮዶን) የሚያካትት የኑክሊክ አሲድ ቅደም ተከተል ገንብተናል። ስለዚህም ሁለቱም ኢንትሮኖች ከ PSC ለጋሽ ተክል ጂን DS. ይህ ቅደም ተከተል በጄንስክሪፕት (በጄንስክሪፕት ባዮቴክ፣ ላይደን፣ ኔዘርላንድስ) ወደ pBINPLUS ተቀላቅሏል። በውስጡ ያለውን ቬክተር እንጠቅሳለን ሲሊ እንደ pBINPLUS-Sli አስገባ።
የ CRISPR ግንባታ-Cas9 ቬክተር. በዲኤም0003 በ PGSC400016861DMG4.03 በ SC እና SI alleles መካከል ምንም አይነት ልዩነት ባልተገኘባቸው ቦታዎች ላይ አራት ጂአርኤንኤን አዘጋጅተናል። ተስማሚ መመሪያዎችን ለመምረጥ እና የቬክተር ግንባታ, በሳንቲላን ማርቲኔዝ እና ሌሎች የተገለጸውን ዘዴ ተጠቀምን.34. ባጭሩ አራት sgRNAs በሊያንግ እና ሌሎች በተገለጹት መመሪያዎች መሰረት ተመርጠዋል።35. የ CC-Top CRISPR/Cas9 ኢላማ ትንበያ መሳሪያ የsgRNAs ዝርዝር ለመፍጠር ጥቅም ላይ ውሏል36፣ ማጠፍ የተገመገመው Mfold ድር አገልጋይን በመጠቀም ነው።37, እና የsgRNAs እንቅስቃሴ የsgRNA መመዝገቢያውን በመጠቀም ተንብዮ ነበር።38. የሚከተሉት መመሪያዎች ተመርጠው ቬክተር pAGM: CRISPRASli: exon5.1T01 (ATTTCATCCGATCTCTCGGGG), exon5.1T04 (GATTTCA TCCGCGATCTCTCGG), exon5.1T06 (TATTTCCTATTGCTACCAGAAGG) እና exon5.1T07 (TATTTCCTATTGCTACCAGAAGG) እና exon86966CGATTTGATTTGATGAT01009CGATTTGATGATTTG0 የ CRISPR ግንባታ ከ Addgene የተገኙትን ፕላዝማይድ በመጠቀም ሠራ: pICH47751 (የማጉላት አብነት); pICSL47761 (ደረጃ 47772 ፕላሲድ); pICH47781, pICH47732, pICH1, pICH41822 እና pICH4723 (ደረጃ 2 plasmids); pICHXNUMX (አገናኝ ለአራት መመሪያዎች); እና pAGMXNUMX (ደረጃ XNUMX ሁለትዮሽ ቬክተር)። ፕላዝማይድ በመጠቀም ክሎክድ ተደርጓል ኢ ኮላይ DH5a እና የተጣራ ፕላዝሚድ በቅደም ተከተል የተላኩት primers PDS5843 (TTTGTGATGCTCGTCAGGGG)፣ PDS8535 (CCCGAGAATTATGCAGCATT TT) PDS8536 (TCATCAGTCAATACATACGGGCT)፣ እና AL717 (GCTTGGCATC 9 Cast መገኘት)፣ የ CastCAGTAGTCATTGGCT፣ እና ALXNUMX (GCTTGGCATC) የአግአACCG መገኘትን ትክክለኛነት ለማረጋገጥ) በመጠቀም ነው።
የpBINPLUS-Sli እና የ pAGMzCRISPR ለውጥAቬክተር ወደ ውስጥ ይንሸራተቱ አግሮባክቲሪየም ቱማፌሲስ. pBINPLUS-Sli ወደ ሆነ ቀይረነዋል አ. tumefaciens AGL0 እና pAGM: CRISPRASliን ወደ ሀ. tumefaciens የኤሌክትሮፖሬሽን ፕሮቶኮልን በመጠቀም AGL0 እና AGL1ን ይሰርዛል። 40 ፒኤል ብቁ የ AGL0 ሴሎችን ወስደን 110 ፕላስ በረዶ-ቀዝቃዛ ሚሊኪው ውሃ ጨምረናል። ከዚህ ድብልቅ 50 ፕላስ ቀድመን ወደተቀዘቀዙ የEppendorf ቱቦዎች በበረዶ ላይ ፓይፕ አድርገን 1 ፒኤል የፕላዝማይድ ጨምረናል። ሴሎቹን በበረዶ ላይ ለ 15 ደቂቃዎች እንተዋቸው እና ሴሎቹን ወደ ቀድሞው ቀዝቃዛ ኤሌክትሮፖሬሽን ኩዌትስ እናስተላልፋለን. ፕሮግራሙን Ec1 (1.8 ኪሎ ቮልት፣ 0.1 ሴሜ ኩቬት) በመጠቀም ውህዶቹን በማይክሮፑልሰር ™ (ባዮ-ራድ ላቦራቶሪዎች፣ ቬ-ኔንዳአል፣ ኔዘርላንድስ) በኤሌክትሮፖሬት አደረግን። 1 ml LB ጨምረናል እና በ 3 ዲግሪ ሴንቲግሬድ እና በ 28 ራፒኤም ላይ ለ 200 ሰዓታት ሴሎቹን ነቅለን. ከዚያ በኋላ፣ Rifampicin (100 pg/ml) እና Kanamycin (50 pg/ml) የያዙ LB agar plates ከትራንስፎርሜሽን ባህል ጋር ገለብን። ሁሉም የተመረጡ ቅኝ ግዛቶች ትክክለኛው ቬክተር እንደያዙ ተረጋግጧል።
የድንች genotypes ለውጥ. ጂኖታይፕን B666 እና B667ን በpBINPLUS-Sli እና genotypes B663 እና B665 ከ pAGM:CRISPRASli ቬክተር ጋር በVisser የተገለጸውን ግንድ ኤክስፕላንት ዘዴን በመጠቀም ቀይረናል።39. ባጭሩ ኢንተርኖድ ኤክስፕላንት የተገኘው በብልቃጥ ውስጥ ከሚበቅሉ ጂኖታይፕስ ሲሆን R3B መካከለኛ ከ2 ሚሊር የPACM መካከለኛ ጋር በፔትሪ ምግቦች ላይ ተቀምጧል። በሚቀጥለው ቀን 50 ሚሊ ሊትር 48 ሰ አግሮባክቲሪየም ባህሎች በ 75 ሚሊ ሊትር LB ውስጥ ማዕከላዊ እና እንደገና ታግደዋል. የ internode explants ከዚያም ውስጥ ሰምጦ ነበር አግሮባክቲሪየም ለ 5 ደቂቃዎች መታገድ ፣ በማጣሪያው ላይ ደርቋል እና እንደገና R3B መካከለኛ በያዙ የፔትሪ ምግቦች ላይ ይቀመጣል። ከ48 ሰአታት ቆይታ በኋላ ገላጣዎቹ ኤምኤስ20ን ወደ ሚይዙት የአንቲባዮቲክ መድኃኒቶች ወደ ፔትሪ ምግቦች ተዘዋውረዋል እና ቡቃያዎችን እንደገና ለማዳበር በእድገት ክፍል ውስጥ እንዲቀመጡ ተደርገዋል። ከታደሰ በኋላ ቡቃያው ሴፎታክሲም (20 ፒጂ/ሚሊ)፣ ቫንኮሚሲን (200 pg/ml) እና ካናማይሲን (200 pg/ml) በያዙ MS100 ሚዲያ ላይ ይበቅላሉ። ቡቃያው በቂ ርዝመት ሲደርስ, ተቆርጦ በ MS20 ውስጥ ያለ አንቲባዮቲክስ ተዘጋጅቷል. በ MS20 ላይ ያለ አንቲባዮቲክስ ቢያንስ ለሁለት ሳምንታት ካደጉ በኋላ ተክሎች በግሪን ሃውስ ውስጥ ተክለዋል.
ፕሎይድ ትንታኔ. የትራንስጀኒክ እፅዋት ፕሎይድ እና ያልተለወጡ መቆጣጠሪያዎች በፕላንት ሳይቶሜትሪ አገልግሎቶች (ዲዳም ፣ ኔዘርላንድ) ፍሰት ሳይቶሜትሪ በመጠቀም ተወስነዋል። ሁሉም የ tetraploid regenerants ተጥለዋል።
የ CRISPR ገጽ ትንታኔ-Cas9-የተፈጠሩ ሚውቴሽን. የዲኤንኤ ማውጣት፣ PCR እና PAGE ትንተና የተካሄደው በሊምግሩፕ (ሆርስት፣ ኔዘርላንድስ) ነው። በCRISPR-Cas9 የታለመውን ክልል ለማጉላት የሚከተሉት ፕሪመርሮች ጥቅም ላይ ውለዋል፡ ወደፊት ፕሪመር፡ CTATTTCCTATTGCTACCAG፣ ተቃራኒ ፕሪመር፡ AAACTTTACCCAAAT AACGTC። የ PCR ምርቶች ስያሜ የተገኘው በግልባጭ ፕሪመር በM13 ጅራት (የመጀመሪያ ቅደም ተከተል፡ TGTAAAACGACGGCCAGTAAACTTTAC CCAAATAAGTC) እና ወይ 700 IRDye ወይም 800 IRDye M13 primer በ PCR ድብልቅ ውስጥ በማከል ነው። የተገኙት PCR ምርቶች በ PAGE ላይ የ Li-cor ስርዓትን በመጠቀም ተንትነዋል። የፒሲአር ምርቶች ከአብዛኛዎቹ መስመሮች ያለ CRISPR-Cas9-induced ሚውቴሽን ከጄል ምስሎች ተወግደዋል Fig. 3d (ኤክሴሽንስ በተሰነጣጠሉ መስመሮች ይገለጻል).
የ 533 bp ማስገቢያ የፋይሎኔቲክ ትንታኔ. የ533 ቢፒ ማስገቢያ ቅደም ተከተል በNCBI ድህረ ገጽ ላይ BLASTn በመጠቀም ተንትኗል። ውስጥ ያለውን ማስገባትን ጨምሮ ምርጡ 30 ምቶች ሲሊ በ Solyntus፣ ከዚያ ወርደው በMegAlign Pro 17 (DNASTAR) MUSCLE ከነባሪ መቼቶች ጋር ተሰልፈዋል። ዛፎቹ የተፈጠሩት በነባሪ ቅንጅቶች የጎረቤት-መቀላቀል አልጎሪዝምን በመጠቀም ነው።
ሪፖርት የማድረግ ማጠቃለያ. ስለ ምርምር ንድፍ ተጨማሪ መረጃ ከዚህ ጽሑፍ ጋር በተገናኘው የተፈጥሮ ምርምር ሪፖርት ማጠቃለያ ውስጥ ይገኛል።
የውሂብ ተገኝነት
የ Solyntus ጂኖም ቅደም ተከተል እና የጥሬ ቅደም ተከተል ንባቦች በNCBI ላይ ይገኛሉ PRJNA631911. የ Solyntus ጂኖም ስብሰባ እና የማብራሪያ ፋይሎች ከWUR ይገኛሉhttps://www.plantbreeding.wur.nl/Solyntus/]. ከበቀለ የአበባ ዱቄት የሚገኘው የአር ኤን ኤ ቅደም ተከተል መረጃ በNCBI አጭር የንባብ መዝገብ ውስጥ ይገኛል PRJNA713577. ሌላ ውሂብ በምንጭ የውሂብ ፋይል ውስጥ ይገኛሉ ወይም ሲጠየቁ ይገኛሉ። የምንጭ መረጃ ከዚህ ወረቀት ጋር ቀርቧል።
የደረሰው: 22 ጥር 2021; ተቀባይነት: 8 ሰኔ 2021;
ማጣቀሻዎች
- 1. ዱቪክ፣ ዲኤን የበቆሎ እድገትን ለማምጣት የመራቢያ አስተዋፅኦ (ዘአ mays ኤል.) Adv. አግሮን። 86፣ 83-145 (2005)።
- 2. Lindhout, P. et al. ወደ F1 ድቅል ዘር ድንች ማራባት። የድንች ሬስ. 54፣ 301-312 (2011)።
- 3. Jansky, SH እና ሌሎች. ድንቹን እንደ ዳይፕሎይድ የተሰራ በመስመር ላይ የተመሰረተ ሰብል በማደስ ላይ። ሳይንስ ይከርክሙ። 56፣ 1412-1422 (2016)።
- 4. ዬ, ኤም. እና ሌሎች. ከ S-RNase በማውጣት ከራስ ጋር የሚስማማ ዳይፕሎይድ ድንች ማመንጨት። ናት. ተክሎች 4፣ 651-654 (2018)።
- 5. Enciso-Rodriguez, F. et al. CRISPR-cas9 ን በመጠቀም በዲፕሎይድ ድንች ውስጥ ራስን አለመጣጣም ማሸነፍ። ግንባር የአትክልት ሳይንስ. 10፣ 1-12 (2019)።
- 6. ሱ, Y. et al. ለመቋቋም የጂኖች መግቢያ ፊቲቶፊቶራ infestans በዲፕሎይድ ድንች ውስጥ. ኤም. ጄ. ድንች ሬስ. 97፣ 33-42 (2020)።
- 7. ዲዚዚንዮ፣ ዲኬ፣ ብራያን፣ ጂጄ፣ ዋይልዴ፣ ጂ እና ሮቢንስ፣ ቲፒ አሌሊክ የ S-RNase alleles በዲፕሎይድ ድንች ዝርያዎች ውስጥ። ተኢዩር. መተግበሪያ ገነት 129፣ 1985-2001 (2016)።
- 8. McClure, B., Cruz-Garcia, F. & Romero, C. በ S-RNase-based ስርዓቶች ውስጥ ተኳሃኝነት እና አለመጣጣም. አን. ቦት 108፣ 647-658 (2011)።
- 9. ኩቦ, K. et al. በS-RNase ውስጥ የትብብር ራስን የማያውቅ ስርዓት - የተመሰረተ ራስን አለመጣጣም። ሳይንስ 330፣ 796-799 (2010)።
- 10. De Jong, H. & Rowe, PR በተመረተ ዳይፕሎይድ ድንች ውስጥ ማዳቀል። የድንች ሬስ. 14፣ 74-83 (1971)።
- 11. ሄርምሰን፣ ጄጂቲ እና ኦልስደር፣ ጄ. ጀነቲክስ ራስን ተኳሃኝነት በዲሃፕሎይድ Solanum tuberosum L. 1. ሁለት ከራስ ጋር የሚጣጣሙ ዲሃፕሎይድስ የመራቢያ ባህሪ. ዩፊቲካ 25፣ 597-607 (1976)።
- 12. ሆሳካ፣ ኬ. እና ሃነማን፣ RE Jr. ከራስ ጋር ተኳሃኝ በሆነው የዱር ዳይፕሎይድ ድንች ዝርያ ውስጥ ያለው ጀነቲክስ Solanum chacoense. 1. የ S locus inhibitor (Sli) ጂን መለየት። ዩፊቲካ 99፣ 191-197 (1998)።
- 13. ሆሳካ፣ ኬ. እና ሃነማን፣ RE Jr. ከራስ ጋር ተኳሃኝ በሆነው የዱር ዳይፕሎይድ ድንች ዝርያ ውስጥ ያለው ጀነቲክስ Solanum chacoense. 2. የዲኤንኤ ምልክቶችን በመጠቀም የ S locus inhibitor (Sli) ጂን በድንች ጂኖም ላይ መፈጠር። ዩፊቲካ 103፣ 265-271 (1998)።
- 14. Birhman, RK & Hosaka, K. የ S-locus inhibitor (Sli) ጂን በመጠቀም የተዳቀሉ የዲፕሎይድ ድንች ዘሮች ማምረት እና ባህሪያቸው። ጂኖም 502፣ 495-502 (2000)።
- 15. Chen, J., Hu, Q., Zhang, Y., Lu, C. & Kuang, H. P-MITE: የተገለበጠ-የሚደጋገሙ ተንቀሳቃሽ ንጥረ ነገሮች የውሂብ ጎታ። ኒውክሊክ አሲድ ችች. 42፣ 1176-1181 (2014)።
- 16. ስቴፋኖቪች፣ ኬ.፣ ላኖ፣ ኤን. እና ቫን ዳምሜ፣ EJM Plant F-box ፕሮቲኖች - በህይወት እና በሞት መካከል ዳኞች። ክሪት ቄስ ተክል ሳይ. 34፣ 523-552 (2015)።
- 17. Liu, B., Morse, D. & Cappadocia, M. Glycosylation of S-RNases በ Solanum chacoense ውስጥ የአበባ ብናኝ ውድቅ ጣራዎችን ሊነካ ይችላል. ጄ. ኤክስፕ. ቦት 59፣ 545-552 (2008)።
- 18. Laimbeer, FPE Potato Genomics በሦስት መንገዶች: በ ሀረጎችና ውስጥ endoreduplication መጠን, transposon መልከዓ ምድር በኩል romp, እና የአበባ ቀለም ደንብ ማብራሪያ. http://hdl.handle.net/10919/84480 (2018).
- 19. ክሎት, ሲአር እና ሌሎች. በድንች ውስጥ በ Sli ላይ የተመሠረተ ራስን መቻል አመጣጥ እና ሰፊ ክስተት። ተኢዩር. መተግበሪያ ገነት https://doi.org/10.1007/s00122-020-03627-8 (2020).
- 20. Endelman, J., Jansky, SH, Butler, N. & Christensen, G. በድንች ክሮሞሶም ላይ ሪሴሲቭ ገዳይ የሆነ የዘረመል ማስረጃ በአሜሪካ የድንች ማህበር የ12-ዓመት ሪፖርት። ኤም. ጄ. ድንች ሬስ. 96, 331 (2019).
- 21. ቫን Lieshout, N. et al. ሶሊንተስ፣ አዲሱ በጣም ተያያዥነት ያለው የድንች ጂኖም (Solanum tuberosum)። G3 ጂኖች ጂኖም ገነት. 10፣ 3489-3495 (2020)።
- 22. Leisner, CP et al. የጂኖም ቅደም ተከተል M6፣ ከፍተኛ-ግላይኮሎይድ የሚያመነጨው የሳንባ ነቀርሳ የድንች ዝርያ ያለው የዲፕሎይድ ክሎሎን Solanum chacoense, ቀሪውን heterozygosity ያሳያል። ተክል ጄ. 1967፣ 562-570 (2018)።
- 23. ፒተርሰን, ቢኤ እና ሌሎች. በራስ መራባት በተመረተ ዳይፕሎይድ ድንች ህዝብ ውስጥ ኢንፊኒየም 8303 ድንች ነጠላ ኑክሊዮታይድ ፖሊሞርፊዝም ድርድር ተመርምሯል። የእፅዋት ጂኖም https://doi.org/10.3835/plantgenome2016.01.0003 (2016).
- 24. ሊያን, Q. et al. ድንች ፖሊፕሎይድ በሚደረግበት ጊዜ ጎጂ ሚውቴሽን ማግኘት። ጄ. ኢንተገር ተክል ባዮ. 61፣ 7-11 (2019)።
- 25. ቫን Lieshout, N. et al. ሶሊንተስ፣ አዲሱ ለድንች በጣም ተያያዥነት ያለው ጂኖም (Solanum tuberosum). G3 ጂኖች ጂኖም ገነት. 631911, g3.401550.2020 (2020).
- 26. ቫን Ooijen, JW ካርታ ይቀላቀሉ®4, በሙከራ ህዝብ ውስጥ የጄኔቲክ ትስስር ካርታዎችን ለማስላት ሶፍትዌር ጥራዝ. 33 (Kyazma BV, 2006)
- 27. ቫን Ooijen, JW በራስ-ጋሞ ዝርያዎች ውስጥ የካርታ መጠናዊ ባህሪ loci ትክክለኛነት። ተኢዩር. አፕል ገነት. 84፣ 803-811 (1992)።
- 28. Voorrips, RE Mapchart: የግንኙነት ካርታዎች እና QTL ስዕላዊ አቀራረብ ሶፍትዌር. ጄ ሄረድ 93፣ 77-78 (2002)።
- 29. ሂርሽ, ሲዲ እና ሌሎች. Spud DB፡ የድንች እርባታን ለማፋጠን የማዕድን ቅደም ተከተሎች፣ ጂኖታይፕስ እና ፍኖታይፕስ ምንጭ። የእፅዋት ጂኖም. 7, https://doi.org/ 10.3835 / plantgenome2013.12.0042 (2014).
- 30. Bankevich, A. et al. SPAdes፡ አዲስ የጂኖም መሰብሰቢያ ስልተ ቀመር እና አፕሊኬሽኑ ወደ ነጠላ ሕዋስ ቅደም ተከተል። ጄ. ኮምፒውተር ባዮ. 19፣ 455-477 (2012)።
- 31. Sharma, SK እና ሌሎች. ለድንች የማጣቀሻ ክሮሞሶም-ልኬት pseudomolecules ግንባታ-የድንች ጂኖምን ከጄኔቲክ እና ፊዚካል ካርታዎች ጋር ማዋሃድ። G3 ጂኖች ጂኖም ገነት. 3፣ 2031-2047 (2013)።
- 32. ፈርናንዴዝ-ፖዞ, N. et al. የ Sol Genomics Network (SGN) - ከጂኖታይፕ እስከ ፍኖታይፕ እስከ እርባታ ድረስ. ኒውክሊክ አሲድ ችች. 43፣ D1036-D1041 (2015)።
- 33. ኪም, ኤስ እና ሌሎች. የሙቅ በርበሬ ጂኖም ቅደም ተከተል በ ውስጥ የፐንጊኒዝም እድገት ላይ ግንዛቤዎችን ይሰጣል Capsicum ዝርያዎች. ናታል. ጀነር. 46፣ 270-278 (2014)።
- 34. Santillan Martinez, MI et al. CRISPR/Cas9-ያነጣጠረ የቲማቲም ተጋላጭነት ጂን PMR4 የዱቄት አረምን ለመቋቋም። BMC ተክል ባዮ. 20፣ 1-13 (2020)።
- 35. Liang, G., Zhang, H., Lou, D. & Yu, D. CRISPR/Cas9 ላይ የተመሰረተ የእጽዋት ጂኖም አርትዖት ከፍተኛ ብቃት ያላቸው sgRNAs ምርጫ። Sci. ሪፐብሊክ. 6፣ 1-8 (2016)።
- 36. Stemmer, M., Thumberger, T., Del Sol Keyer, M., Wittbrodt, J. & Mateo, JL CCtop: ሊታወቅ የሚችል, ተለዋዋጭ እና አስተማማኝ CRISPR/Cas9 የዒላማ ትንበያ መሳሪያ. PLoS ONE 10,1-11 (2015).
- 37. Zuker, M. Mfold ዌብ አገልጋይ ለኑክሊክ አሲድ መታጠፍ እና ማዳቀል ትንበያ። ኒውክሊክ አሲድ ችች. 31፣ 3406-3415 (2003)።
- 38. Chari, R., Yeo, NC, Chavez, A. & Church, GM SgRNA Scorer 2.0: የ CRISPR/Cas9 እንቅስቃሴን ለመተንበይ ከዝርያ-ገለልተኛ ሞዴል. ኤሲኤስ ሲንት ባዮ. 6፣ 902-904 (2017)።
- 39. Visser, RGF ውስጥ የእፅዋት ቲሹ ባህል መመሪያ 301-309 (ስፕሪንግ, 1991).
ተወዳጅ ፍላጎቶች
ደራሲዎቹ ምንም የተወዳጅ ፍላጎት አይኖራቸውም.
ተጭማሪ መረጃ
ተጨማሪ መረጃ የመስመር ላይ እትሙ በ ላይ የሚገኘውን ተጨማሪ ነገር ይዟል https://doi.org/10.1038/s41467-021-24267-6.
የተመጣጠነ መሆን እና የቁሳቁስ ጥያቄዎች ወደ CWBB መቅረብ አለባቸው
የአቻ ግምገማ መረጃ ተፈጥሮ ግንኙነቶች ሮጀር ቼቴላትን እና ሌላውን ስም የለሽ ገምጋሚ(ዎች) ለዚህ ስራ የአቻ ግምገማ ላደረጉት አስተዋፅዖ እናመሰግናለን። የአቻ ግምገማ ሪፖርቶች ይገኛሉ።
የድጋሚ ህትመት እና የፍቃድ መረጃ በ የሚገኘው ይገኛል http://www.nature.com/reprints
አታሚ's ማስታወሻ Springer Nature በተፈቀዱ ካርታዎች እና በተቋማዊ ተያያዥነት ላይ በሚነሱ የአስተዳደር ጉዳዮች ላይ ገለልተኛ ነው.
ማረጋገጫዎች
ከቬሮኒካ ታሚ ሶፑትሮ የተገኘውን ሁለት የድንች ጂኖቲፕስ ለውጥ በ pAGM:CRISPRASli፣ የቢኤስሲ እና የኤምኤስሲ ተማሪዎች ጊሎ ፕሊኒስ፣ አይሪስ ስሚትስ፣ ንጉሴ ቮርጊያ፣ ሂድድ ክኑይማን፣ ሞሪስ ጊውርትስ፣ አንጃ ቫን ሄተርን እና ቶርስተን ቫን ደር እገዛ እውቅና እንሰጣለን። ሼሪክ የካርታውን ህዝብ በፍኖተ ካርታ በመፃፍ እና ቴስ ሉካስ የአር ኤን ኤ ናሙናዎችን በማመንጨት ለረዳቸው። በተጨማሪም የ Solynta እና Wageningen University Unifarm ግሪንሃውስ ሰራተኞች በእጽዋት እንክብካቤ እና በዘር ማውጣት ላይ ላደረጉት እገዛ እናመሰግናለን። ይህ ፕሮጀክት ከኔዘርላንድ የሳይንቲፊክ ምርምር ድርጅት የገንዘብ ድጋፍ አግኝቷል (የስጦታ መታወቂያ፡ NWA.17.023)።
የደራሲ አስተዋጽኦዎች
E.-JE ሙከራዎቹን ነድፎ ፈጽሟል እና የእጅ ጽሑፉን ጻፈ። AvdB የWGS፣ KASP ማርከርን እና የባዮኢንፎርማቲክስ አቀራረቦችን ነድፎ ፈጽሟል። SvH፣ RGFV፣ CWBB፣ MEDV፣ እና PL ለጄኔቲክ ካርታ ጥናት እና ተግባራዊ ባህሪ አቀራረቡን ለመንደፍ ረድተዋል። ሲሊ. RGFV፣ CWBB እና PL የእጅ ጽሑፉን ገምግመው አስተያየት ሰጥተዋል።
I(S) ጥ) መዳረሻ ክፈት ይህ አንቀፅ ለዋናው ደራሲ (ቶች) እና ለነጩ ምንጭ ተገቢውን ብድር እስከሰጡ ድረስ ፣ በየትኛውም መካከለኛ ወይም ቅርጸት መጠቀም ፣ ማጋራት ፣ መላመድ ፣ ማሰራጨት እና ማራባት በሚፈጥረው በፈረንሳዊው የጋራ ባህሪዎች 4.0 ዓለም አቀፍ ፈቃድ ስር ፈቃድ የተሰጠው ነው ፡፡ ወደ Creative Commons ፈቃዱ አገናኝ ያገናኙ እና ለውጦች እንደተደረጉ ያመልክቱ። በዚህ ጽሑፍ ውስጥ ምስሎቹ ወይም ሌሎች የሦስተኛ ወገን ይዘቶች በቁሳቁስ ላይ በዱቤ መስመር ካልተገለፀ በቀር በአንቀጹ የፈጠራ ጋራዎች ፈቃድ ውስጥ ተካትተዋል ፡፡ ይዘቱ በጽሁፉ የፈጠራ ጋራዎች ፈቃድ ውስጥ ካልተካተተ እና የታሰበው የእርስዎ ጥቅም በሕጋዊ ደንብ ካልተፈቀደ ወይም ከሚፈቀደው አገልግሎት የሚልቅ ከሆነ በቀጥታ ከቅጂ መብት ባለቤቱ በቀጥታ ፈቃድ ማግኘት ያስፈልግዎታል። የዚህን ፈቃድ ቅጂ ለማየት ፣ ይጎብኙ http://creativecommons.org/ ፍቃዶች/በ/4.0/.





